
下载掌阅APP,畅读海量书库
立即打开



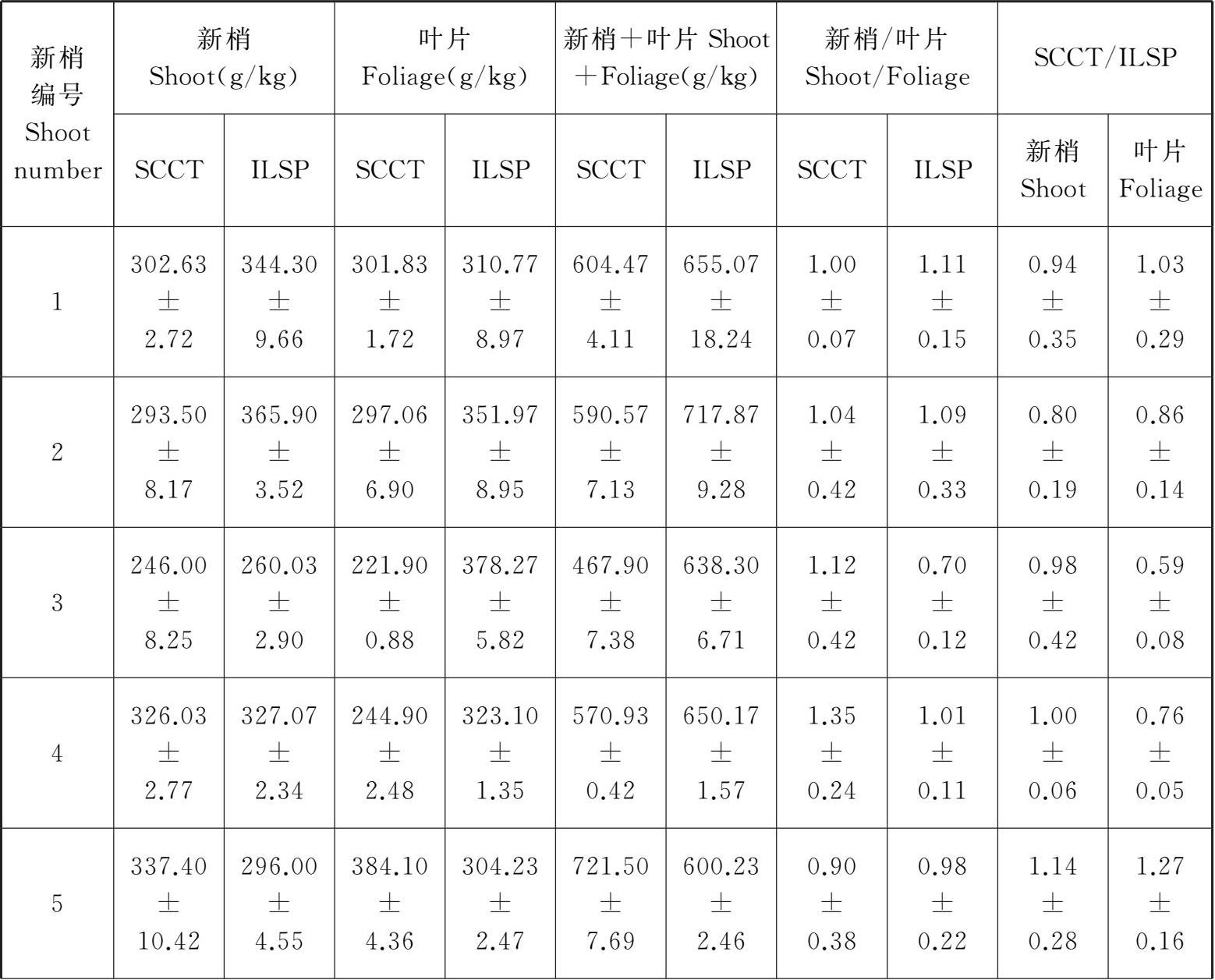
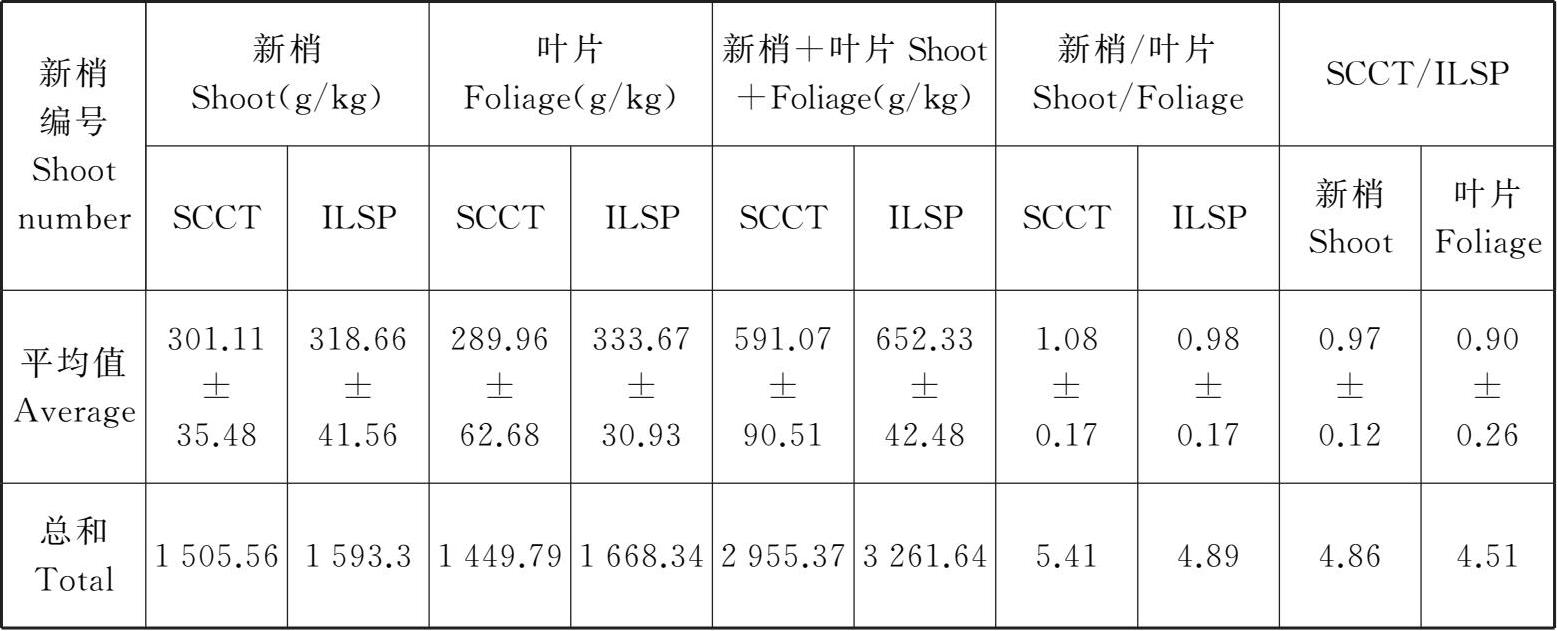
从表3-1可以看出,在10月上旬,随新梢部位的变化,SCCT和ILSP的5个新梢或叶片的有机碳积累总量依次为 1 505.56 g/kg、1 593.3 g/kg、1 449.79 g/kg和1 668.34 g/kg,而SCCT和ILSP的 5 个新梢和叶片的有机碳积累总量依次为2 955.37 g/kg和3 261.64 g/kg,SCCT和ILSP的5个新梢和叶片的有机碳积累总量之比依次为5.41和4.89,五个新梢或叶片在SCCT和ILSP两种架式下的有机碳积累总量比值依次为4.86和4.51。
表3-1 爱格丽葡萄新梢和叶片有机碳含量
续表
注:表中数据为平均值±标准差。下同。
1、2、3、4和5分别表示同一葡萄植株从基部开始的5个新梢或相应叶片的编号。每一种架式的新梢或相应叶片的有机碳含量为该新梢的上、中和下三个部位有机碳含量的平均值,其值反映了有机碳含量在该新梢或相应叶片上的分配状况;每一种新梢上叶片的有机碳含量为该新梢上、中和下三个部位叶片的有机碳含量的平均值,其值反映了有机碳含量在对应新梢上的叶片的分配状况;新梢+叶片表示同一植株每一种架式的地上部分各器官(新梢及相应叶片)有机碳含量的总和,其值反映了该架式的葡萄植株地上部分的固碳能力;新梢/叶片表示每一种架式的新梢与相应叶片的有机碳含量之比,其值反映了有机碳含量在该新梢与相应叶片的分配比例;SCCT/ILSP分别表示SCCT与ILSP的新梢或叶片的有机碳含量之比,其值反映了两种架式对同一株葡萄新梢或叶片中有机碳含量分配的影响。这与架式影响环境中有机碳的可利用性以及植物对有机碳的吸收和运输能力有关。
10月上旬,不同架式的新梢和叶片中有机碳含量存在较大的差别。就新梢而言,SCCT的5个新梢有机碳积累量平均为301.11 g/kg,而ILSP的5个新梢有机碳积累量平均为318.66 g/kg;就叶片而言,SCCT的5个新梢相应叶片的有机碳积累量平均为289.96 g/kg,而ILSP的5个新梢相应叶片的有机碳积累量平均为333.67 g/kg。综合比较,同一器官不同架式间的有机碳积累量差异明显,以SCCT叶片的有机碳含量最低(221.9 g/kg),ILSP叶片的有机碳含量最高(378.27 g/kg);各器官的有机碳含量随着部位而变化,SCCT叶片和新梢的有机碳含量均是从基部第一个新梢开始降低,在第三个新梢降到最低,之后开始升高;而ILSP的5个新梢量的有机碳含量呈“M”型变化,SCCT的5个新梢量的有机碳含量呈“Λ”型变化。
比较10 月上旬SCCT和ILSP两种架式的爱格丽五个新梢有机碳含量发现(表3-1),除了第五个新梢外,爱格丽葡萄的SCCT的其余新梢的有机碳含量明显低于ILSP新梢的有机碳含量;SCCT新梢的有机碳含量中,第五个新梢的有机碳含量最高,为337.40 g/kg,占整个植株新梢有机碳含量的22.41%,其次依次为第四个新梢、第一个新梢、第二个新梢和第三个新梢。ILSP各新梢的有机碳含量中,第二个新梢的有机碳含量最高,为365.90 g/kg,占整个植株新梢有机碳含量的22.96%,其次依次为第一个新梢、第四个新梢、第五个新梢和第三个新梢的有机碳含量。两种架式的第三个新梢的有机碳含量均为最低,但是ILSP第三个新梢的有机碳含量仍大于SCCT第三个新梢的有机碳含量。这与架式影响环境中有机碳的可利用性以及植物对有机碳的吸收和运输能力有关。
比较10月上旬SCCT和ILSP两种架式的爱格丽葡萄五个新梢叶片的有机碳含量发现(表3-1),除了第五个新梢上的叶片外,SCCT其余新梢上的叶片的有机碳含量明显低于ILSP新梢上的叶片的有机碳含量。SCCT各新梢上的叶片的有机碳含量中,第五个新梢上的叶片的有机碳含量最高,为384.10 g/kg,占整个植株新梢上叶片有机碳含量的26.49%,其次依次为第一个新梢、第二个新梢、第四个新梢和第三个新梢上的叶片。ILSP各新梢上的叶片的有机碳含量中,第三个新梢上叶片的有机碳含量最高,为378.27 g/kg,占整个植株新梢上的叶片有机碳含量的22.67%,其次依次为第二个新梢、第四个新梢、第一个新梢和第五个新梢上的叶片。SCCT第三个新梢上的叶片的有机碳含量为最低,而ILSP第五个新梢上的叶片的有机碳含量为最低,但是ILSP第五个新梢上的叶片的有机碳含量仍大于SCCT第三个新梢上的叶片的有机碳含量。这与架式影响环境中有机碳的可利用性以及植物对有机碳的吸收和运输能力有关。
比较10月上旬SCCT和ILSP两种架式的爱格丽葡萄的地上部分各器官(新梢+叶片)有机碳含量发现(表3-1),与新梢或叶片相似,除了第五个新梢+叶片外,SCCT的其余新梢+叶片的有机碳含量明显低于ILSP新梢+叶片的有机碳含量。SCCT各新梢+叶片的有机碳含量中,第五个新梢+叶片的有机碳含量最高,为721.50 g/kg,占整个植株新梢+叶片有机碳含量的24.41%,其次依次为第一个新梢+叶片、第二个新梢+叶片、第四个新梢+叶片和第三个新梢+叶片。ILSP各新梢+叶片的有机碳含量中,第二个新梢+叶片的有机碳含量最高,为717.87 g/kg,占整个植株新梢+叶片有机碳含量的22.01%,其次依次为第一个新梢+叶片、第四个新梢+叶片、第三个新梢+叶片和第五个新梢+叶片的有机碳含量。SCCT第三个新梢+叶片的有机碳含量为最低,而ILSP第五个新梢+叶片的有机碳含量为最低,但是ILSP第五个新梢+叶片的有机碳含量仍大于SCCT第三个新梢+叶片的有机碳含量。这与架式影响环境中有机碳的可利用性以及植物对有机碳的吸收和运输能力有关。
比较10月上旬SCCT和ILSP两种架式的爱格丽葡萄五个新梢与叶片的有机碳含量比值发现(表3-1),除了SCCT的第四个和第三个新梢与叶片的有机碳含量的比值大于相应的ILSP新梢与叶片的有机碳含量比值外,爱格丽葡萄的SCCT的其余三个新梢与叶片的有机碳含量比值明显低于ILSP新梢的有机碳含量比值。SCCT各新梢与叶片的有机碳含量比值中,第四个新梢与叶片的有机碳含量比值最高,为1.35,其次依次为第三个新梢、第二个新梢、第一个新梢和第五个新梢与叶片的有机碳含量比值。ILSP各新梢与叶片的有机碳含量比值中,第一个新梢的有机碳含量最高,为1.11,其次依次为第二个新梢、第四个新梢、第五个新梢和第三个新梢与叶片的有机碳含量比值。SCCT第五个新梢与叶片的有机碳含量比值为最低,而ILSP第三个新梢与叶片的有机碳含量比值最低,而且SCCT第五个新梢与叶片的有机碳含量比值仍大于ILSP第三个新梢与叶片的有机碳含量比值。这与架式影响环境中有机碳的可利用性以及植物对有机碳的吸收和运输能力有关。
分别比较10月上旬爱格丽葡萄的五个新梢或叶片在SCCT和ILSP两种架式下的有机碳含量比值发现(表3-1),除了第四个和第三个新梢在SCCT和ILSP两种架式下的有机碳含量比值大于相应叶片在SCCT和ILSP两种架式下的有机碳含量比值外,爱格丽葡萄的其他部位新梢在SCCT和ILSP两种架式下的有机碳含量比值小于相应叶片在SCCT和ILSP两种架式下的有机碳含量比值。不同部位新梢在SCCT和ILSP两种架式下的有机碳含量比值中,第五个新梢在SCCT和ILSP两种架式下的有机碳含量比值最高,为1.14,其次依次为第四个新梢、第三个新梢、第一个新梢、第二个新梢在SCCT和ILSP两种架式下的有机碳含量比值。不同部位新梢对应的叶片在SCCT和ILSP两种架式下的有机碳含量比值中,第五个新梢对应的叶片在SCCT和ILSP两种架式下的有机碳含量比值最高,为1.27,其次依次为第一个新梢、第二个新梢、第四个新梢和第三个新梢对应的叶片在SCCT和ILSP两种架式下的有机碳含量比值。第二个新梢在SCCT和ILSP两种架式下的有机碳含量比值最低,而第三个新梢对应的叶片在SCCT和ILSP两种架式下的有机碳含量比值最低,而且第二个新梢在SCCT和ILSP两种架式下的有机碳含量比值仍大于第三个新梢对应的叶片在SCCT和ILSP两种架式下的有机碳含量比值。这与架式影响环境中有机碳的可利用性以及植物对有机碳的吸收和运输能力有关。
通常,新梢与叶片的有机碳含量比值大于1时,表明新梢中吸收或贮存的有机碳含量大于叶片吸收或贮存的有机碳含量;值越大,新梢中吸收或贮存的有机碳含量比叶片吸收或贮存的有机碳含量多;反之亦然。新梢与叶片的有机碳含量比值小于1时,表明新梢中吸收或贮存的有机碳含量小于叶片吸收或贮存的有机碳含量;值越小,新梢中吸收或贮存的有机碳含量比叶片吸收或贮存的有机碳含量少;反之亦然。新梢与叶片的有机碳含量比值等于1,新梢中吸收或贮存的有机碳含量与叶片吸收或贮存的有机碳含量相当。这个比值在植株不同生长期反映了植株不同器官对有机碳含量的吸收或贮存状况,也反映了植株所处的不同生长期及植株的成熟状况。
同理,不同处理的植株某一器官的有机碳含量比值大于1时,这种处理对某一器官吸收或贮存有机碳含量的能力占优势;值越大,这种处理对某一器官吸收或贮存有机碳含量的能力越优越;这种处理的某一器官吸收或贮存有机碳的含量越多;反之亦然。不同处理的植株某一器官的有机碳含量比值小于1时,表明这种处理对某一器官吸收或贮存有机碳含量的能力不占优势;值越小,这种处理对某一器官吸收或贮存有机碳含量的能力越不明显;这种处理的某一器官吸收或贮存有机碳的含量越少;反之亦然。不同处理的植株某一器官的有机碳含量比值等于1时,这种处理对某一器官吸收或贮存有机碳含量的能力相当,采用何种处理都不会影响某一器官对有机碳含量的吸收或贮存。这个比值在植株不同生长期反映了不同处理对植株某一器官有机碳含量的吸收或贮存能力的影响,也反映了这一处理对植株在不同生长期的成熟状况的影响。
植物的形态结构和生理机制对环境的适应,能使植物最有效地利用环境资源,从而趋于最佳生长状态 [23] 。架式影响了植物的生长环境和生长状况,所以,影响着葡萄植株新梢和叶片有机碳含量的分配特征。SCCT与ILSP两种架式的架型不同,前者的架型较矮,且主干高度极短,一条平行生长的龙干上着生的新梢垂直于龙干生长,而后者的架型较高,且主干较长,一条倾斜于地面的主干上着生的新梢在距地面50 cm的高度呈“V”型生长。植物器官本身的结构和功能差异影响着营养元素在植物器官中的分配。所以,两种架式的葡萄植株的新梢和叶片中,有机碳含量都是ILSP的最高(新梢为365.90 g/kg,叶片为 378.27 g/kg),SCCT的最低(新梢为 246.00 g/kg,叶片为221.90 g/kg)。SCCT新梢和叶片中的有机碳含量多低于ILSP相应的新梢和叶片中的有机碳含量。通常,植物在生长期会将大部分营养元素转移到地上部分各器官中 [25] ,不同组织器官中的营养元素分配为叶中最大,其次为新梢 [26] ,本研究结论与之不完全一致。叶作为光合作用的主要器官,其光合产物主要集中在叶肉细胞中,因此,葡萄叶片中的有机碳含量较高是正常的。由于本实验中的样品采于10月上旬,植物的生理活动主要表现贮存养分,一方面,叶片的光合产物由叶片向新梢和根系转移,叶片吸收和固定碳的能力减弱;另一方面,植物根系从土壤获得的有机碳向新梢转移,以促进新梢的木质化,导致了新梢中积累或贮存了大量的有机碳。新梢中积累或贮存的有机碳含量大于叶片中积累或贮存的有机碳含量(如SCCT第四枝和ILSP的头两枝),或者新梢中积累或贮存的有机碳含量与叶片中积累或贮存的有机碳含量持平(表3-1)。叶片中的有机碳含量高于新梢中的有机碳含量(SCCT第五枝与ILSP第三枝和第五枝),主要是由于此类叶片仍处于光合作用的高峰期,其吸收和固定碳的能力较强,同时又由于叶片不断向新梢和根系转移碳,所以,叶片中固定的有机碳和相应新梢转移的有机碳含量均高,而且叶片中固定的有机碳仍然高于叶片向相应新梢中转移的有机碳(如SCCT第五枝)(表3-1),这也是同一植株不同部位新梢和相应叶片有机碳含量不一致的原因之一。
架式引起了葡萄叶面积指数的变化,进而引起了的净光合速率、气孔导度和蒸腾速率等指标的改变 [27-28] ,从而影响了植物对养分的运输。
本实验中,对于两种架式的新梢和叶片的有机碳含量来说,除了第五个新梢和叶片外,SCCT其他新梢和叶片的有机碳含量均少于相应的ILSP新梢和叶片的有机碳含量。由于SCCT五个新梢直立的架式,其生长期每个新梢的叶片接受光照的面积比ILSP大,每个新梢的叶片接受的光照强度比ILSP强,所以SCCT的叶片比ILSP的叶片成熟早,SCCT的新梢比ILSP木质化快,SCCT叶片中的有机碳大部分通过新梢转移到了根系或龙干中贮存起来。SCCT的叶片和相应的新梢有机碳含量分配较均衡;而在生长期,ILSP的五个新梢倾斜向上直立生长,其每个叶片接受光照的面积比SCCT小且不均匀,每个叶片接受的光照强度比SCCT弱且不一致,所以ILSP的叶片比SCCT的叶片成熟晚且不均衡,ILSP的新梢比SCCT木质化慢且不协调,导致了ILSP的叶片和相应的新梢有机碳含量分配不均衡。同时,由于ILSP的叶片成熟较晚,此时其吸收和固定有机碳的能力仍较强,所以,其叶片的有机碳含量比SCCT丰富,其叶片运输或转移到相应新梢上的有机碳含量也比SCCT丰富。因此,ILSP新梢和叶片内的有机碳含量比SCCT相应新梢和叶片内的有机碳含量多,且ILSP新梢和叶片内的有机碳含量分配不均衡。
活有机体的元素组成相对比较稳定,与植物细胞自身的缓冲性有关 [29] 。所以,不同架式的同一植物体内生长期碳元素含量之间存在相对稳定的比值关系,不同架式的新梢和叶片碳元素间比例关系的变化可以反映植物体生长的状况,进一步反映植物体不同器官的营养状况和碳元素分配供应情况。两种架式葡萄的生长环境(主要是水分供应、光照和温度)的差异影响营养元素碳的可利用性 [30] 以及植物对养分的吸收 [31] 和运输,进而导致不同架式不同器官间碳碳比的差异。不同生长时期,植物各器官碳碳比的变化均具有时间性。在生长末期,植物不同部位的叶片光合作用发生了变化,植物下部的叶片因为过早衰败而导致了光合作用效率降低,上部叶片因为生长较旺盛,光合速率较高而具有较高的有机碳含量,中部叶片因为光合产物有机碳含量不但供给自己进行生理活动的需要,还要向植株其他部位输送,所以其有机碳含量最低(表3 1)。这与不同时期各器官对碳在各器官的转移和分配状况有关 [32-33] 。生长末期,从基部开始,ILSP新梢与叶片的碳碳比总体上呈现下降趋势,而SCCT新梢与叶片的碳碳比总体上呈现上升趋势,但是第五个新梢与叶片的碳碳比降低了,这与架式的变化有关。
10月上旬爱格丽葡萄两种架式不同器官的有机碳含量平均值见表3 1。两种架式的葡萄新梢和叶片的有机碳都表现为新梢大于叶片,这是植物对养分的一种分配策略。在本实验中,第一至第四个新梢的有机碳含量大于相应叶片的有机碳含量。叶片作为植物同化作用的主要器官,新陈代谢旺盛,植物将养分更多输送到叶片中,以满足快速生长的需求 [34] 。但是在采收后、生长末期或者在新梢木质化过程中,叶片中同化的有机碳不断向新梢转移,促进新梢的木质化和成熟,直至新梢和叶片中的有机碳含量达到平衡。第一至第四个新梢和叶片的碳碳比大于1(除了ILSP的第三个新梢和叶片的碳碳比小于1),而第五个新梢和叶片的碳碳比小于1。等于1,新梢和叶片中的有机碳含量达到平衡;大于1,表明叶片向新梢转移的有机碳含量超过了叶片自身固定的有机碳含量,或处于成熟期,新梢成熟较好,而且,值越大,新梢成熟度越好;小于1,表明叶片向新梢转移的有机碳含量低于叶片自身固定的有机碳含量,新梢还未成熟,处于生长期,而且,值越小,新梢成熟度越差。通过这种方式,也可以间接判断植株或新梢的生长阶段,或者同一植株不同部位新梢或叶片的生长状况,有利于对植株采取正确的管理措施。SCCT第三和第四个新梢和叶片的碳碳比高于ILSP新梢和叶片的碳碳比,而SCCT第一、第二和第五新梢和叶片的碳碳比低于ILSP新梢和叶片的碳碳比,表明SCCT第三和第四个新梢和叶片的成熟比ILSP的好,SCCT第一、第二和第五个新梢和叶片的成熟没有ILSP的好,但是第五个新梢和叶片的碳碳比小于1,表明其成熟度最差。按照新梢和叶片碳碳比的大小,可以将SCCT五个新梢的成熟程度排列为:第四、第三、第二、第一和第五个新梢,其碳碳比的变化围在0.90~1.35,反映了SCCT的同一个植株的五个新梢的成熟程度;ILSP五个新梢的成熟程度排列为:第一、第二、第四、第五和第三个新梢,其碳碳比的变化围在0.70~1.11,反映了ILSP的同一个植株的五个新梢的成熟程度。同理,按照SCCT和ILSP新梢的有机碳之比,可以将五个新梢的成熟程度排列为:第五、第四、第三、第一和第二个新梢,其碳碳比的变化围在0.80~1.14,反映了SCCT的五个新梢分别比ILSP相应的五个新梢的成熟程度;按照SCCT和ILSP新梢上的叶片的有机碳之比,可以将五个新梢上的叶片的成熟程度排列为:第五、第一、第二、第四和第三个新梢上的叶片,其碳碳比的变化围在0.59~1.27,反映了SCCT的五个新梢上的叶片分别比ILSP相应的五个新梢上的叶片的成熟程度。综合分析,每个架式的葡萄树留4个新梢较为合适。
碳是植物体中含量最高的元素之一,它源于植物通过光合作用固定大气中的二氧化碳,并以有机物形式作为能量贮存起来。植物体的碳含量平均为45%左右,主要以糖类、氨基酸、蛋白质、脂肪等有机物的形式存在,植物碳含量高低受品种、器官、生长期以及生境条件等因素的影响,探讨葡萄生长过程中碳含量的动态变化可为全面了解有机物在葡萄中的转运和分配提供科学借鉴,这对维持葡萄植株不同器官的碳平衡、推动葡萄植株不同器官间的物质循环和能量流动至关重要。
碳是植物体的基本框架元素,葡萄成熟过程中积累的碳在各个器官的转运与分配对葡萄新梢和叶片的成熟影响很大。本实验对葡萄不同器官有机碳含量的初步研究表明,碳在葡萄植株不同器官中的分配受架式和物候期的影响。葡萄地上部分不同器官积累有机碳的能力存在差异,这主要与架式导致的葡萄不同器官的功能差异有关。葡萄主要的光合器官是叶片,葡萄叶片光合同化的有机碳通过碳代谢和转运过程分配到各个器官。在葡萄叶片自然衰老和新梢自然休眠或木质化过程中,由于其生理功能衰退,叶片中的有机碳输出,必然造成叶片中有机碳含量的下降,所以,造成了同一植株不同部位有机碳含量的差异,这与Yang et al. [34] 和Lin et al. [35] 对水稻叶片的研究得到的结论一致。架式引起了葡萄植株结构的差异,必然导致葡萄植株不同部位叶片光照的差异,进而引起叶片光合有机碳分配的差异及叶片自然衰老程度的差异。光照充分,叶片成熟快且早,反之亦然。这与物候期也有关系。所以,本实验中,SCCT叶片中的有机碳含量比ILSP叶片中的少。新梢中也得到了相似的结果,主要是SCCT新梢中的有机碳较快转移到了根部贮存起来。但是,SCCT第五个新梢的叶片中的有机碳含量比ILSP的多,主要是由于第五个新梢距离根部太远,叶片同化碳合成、积累、运输和分配较慢,进而导致了SCCT相应新梢的有机碳含量比ILSP的多。所以,架式及架式引起的光照变化影响了葡萄新梢和叶片的有机碳运输与分配,这与Lin et al. [35] 在不同水稻品种上的研究结论一致。进一步的研究应在架式引起的物候期变化,或者在不同的物候期,架式对葡萄植株不同部位有机碳的影响。
因此,两种架式的葡萄新梢和叶片的有机碳含量存在着显著的架式和器官间差异。10月上旬,ILSP的第一、第二、第三和第四个新梢和叶片的有机碳含量均分别大于SCCT新梢和叶片的有机碳含量,而ILSP的第五个新梢和叶片的有机碳含量分别小于SCCT新梢和叶片的有机碳含量。但是SCCT第一、第二、第三和第四个新梢成熟较好,而ILSP第一、第二和第四个新梢成熟较好。总体上,ILSP新梢和叶片积累的有机碳含量高于SCCT新梢和叶片积累的有机碳含量,但是SCCT新梢的成熟度比ILSP好,这与两种架式对葡萄生长环境的影响有关。
[1] Dannenberg S,Conrad R.Effect of rice plants on methane production and rhizospheric metabolism in paddy soil[J]. Biogeochemistry ,1999,45(1):53-71.
[2]Black T A,Chen J M,Lee X,Sagar R M.Characteristics of shortwave and longwave irradiances under a Douglas-fir forest stand[J]. Canadian Journal of Forest Research ,1991,21(7):1020 1028.
[3]Chen J M,Leblanc S G,Miller J R,Freemantle J,Loechel S E,Walthall C L,White H P.Compact airborne spectrographic imager(CASI)used for mapping biophysical parameters of boreal forests[J]. Journal ofGeophysical Research : Atmospheres ,1999,104(D22):27945 -27958.
[4]Friedl M A,McIver D K,Hodges J C,Zhang X Y,Muchoney D,Strahler A H,Schaaf C.Global land cover mapping from MODIS:algorithms and early results[J]. Remote Sensing of Environment ,2002,83(1):287-302.
[5]Han W,Fang J,Guo D,Zhang Y.Leaf nitrogen and phosphorus stoichiometry across terrestrial plant species in China[J]. New Phytologist ,2005,168(2):377-385.
[6]任书杰,于贵瑞,陶波,王绍强.中国东部南北样带654种植物叶片氮和磷的化学计量学特征研究[J].环境科学,2007,28(12):2665-2673.
[7]刘大勇,陈平,范志平,于占源.氮、磷添加对半干旱沙质草地植被养分动态的影响[J].生态学杂志,2006,6:612-616.
[8]林瑞余,蔡碧琼,柯庆明,蔡向阳,林文雄.不同水稻品种产量形成过程的固碳特性研究[J].中国农业科学,2006,12:2441-2448.
[9]展茗.不同稻作模式稻田碳固定、碳排放和土壤有机碳变化机制研究[D].武昌:华中农业大学,2009.
[10]吴志丹,王义祥,翁伯琦,蔡子坚,温寿星.福州地区7年生柑橘果园生态系统的碳氮储量[J].福建农林大学学报(自然科学版),2008,37(3):316-311.
[11]王月.CO 2 浓度升高对不同供磷番茄根系生长和根系分泌物的影响[D].杭州:浙江大学,2008.
[12]刘雪云,周志宇,郭霞,王瑞,梁坤伦.紫穗槐植株的养分含量及分布特征[J].草业学报,2012,5:264-273.
[13]Grossman Y L,DeJong T M.Training and pruning system effects on vegetative growth potential,light interception,and cropping efficiency in peach trees[J]. Journalofthe American Societyfor Horticultural Science ,1998,123(6):1058 -1064.
[14]Robertson G P,Paul E A, Harwood R R.Greenhouse gases in intensive agriculture:contributions of individual gases to the radiative forcing of the atmosphere[J]. Science ,2000,289(5486):1922-1925.
[15]Janssens I A,Freibauer A,Ciais P,Smith P,Nabuurs G J,Folberth G,Dolman A J.Europe’s terrestrial biosphere absorbs 7 to 12% of European anthropogenic CO 2 emissions[J]. Science ,2003,300(5625):1538-1542.
[16]Proctor J T A,Watson R L,Landsberg J J.The carbon budget of a young apple tree[J]. Journal of the American Societyfor Horticultural Science ,1976,101:579-582.
[17]Ebert G,Lenz F.Annual course of root respiration of apple trees and its contribution to the CO 2 -balance[J]. Gartenbauwissenschaft ,1991,56:130-133.
[18]Wibbe M L,Blanke M M,Lenz F.Effect of fruiting on carbon budgets of apple tree canopies[J]. Trees ,1993,8(1):56-60.
[19]Blanke M M.Contribution of soil respiration to the carbon balance of an apple orchard[J].In VI International Symposium on Integrated Canopy,Rootstock,Environmental Physiology in Orchard Systems,1996,451:337-344.
[20]Sekikawa S,Kibe T,Koizumi H,Mariko S.Soil carbon budget in peach orchard ecosystem in Japan[J]. Environmental Science ,2003,16(2):97-104.
[21]Sofo A,Nuzzo V,Palese A M,Xiloyannis C,Celano G,Zukowskyj P,Dichio B.Net CO 2 storage in mediterranean olive and peach orchards[J]. Scientia horticulturae ,2005,107(1):17-24.
[22]那守海,郝铁钢,阎秀峰.供氮水平对落叶松根系碳、氮积累与分配的影响[J].东北林业大学学报,2007,11:17-19+22.
[23]Pessarakli M.Handbook of photosynthesis[M].2nd(Ed).London:CRC Press,2005,169 451.
[24]管东生,罗琳.海南热带植物叶片化学元素含量特征[J].林业科学,2003,39(2):28-32.
[25]莫江明,张德强,黄忠良,余清发,孔国辉.鼎湖山南亚热带常绿阔叶林植物营养元素含量分配格局研究[J].热带亚热带植物学报,2000,3:198-206.
[26]张大鹏,娄成后.北京地区葡萄三种主要栽培方式的叶幕微气候和植物水分关系的研究[J].中国农业科学,1990,23(02):73-82.
[27]Iacono F,Sommer K J.Photoinhibition of photosynthesis and photorespiration in Vitis vinifera under field conditions-effects of light climate and leaf position[J]. Australian Journal ofGrape and Wine Research ,1996,2(1):1 -11.
[28]曾德慧,陈广生.生态化学计量学:复杂生命系统奥妙的探索[J].植物生态学报,2005,29(6):1007-1019.
[29]杨继松,刘景双,于君宝,王金达,李新华,孙志高.三江平原沼泽湿地枯落物分解及其营养动态[J].生态学报,2006,26(5):1297-1302.
[30]Rubio G,Lavado R S.Acquisition and allocation of resources in two waterlogging-tolerant grasses[J]. New Phytologist ,1999,143(3):539-546.
[31]孙志高,刘景双,王金达,秦胜金.三江平原不同群落小叶章种群生物量及氮、磷营养结构动态[J].应用生态学报,2006,2:221-228.
[32]孙志高,刘景双,于君宝.三江平原不同群落小叶章氮素的累积与分配[J].应用生态学报,2009,2:277-284.
[33]郑淑霞,上官周平.黄土高原地区植物叶片养分组成的空间分布格局[J].自然科学进展,2006,16(8):965-973.
[34]Yang J C,Zhang J H,Wang Z Q,Zhu Q S,Wang W.Remobilization of carbon reserves in response to water deficit during grain filling of rice[J]. Field Crops Research ,2001,71(1):47-55.
[35]Lin X Q,Wang Y F,Zhu D F,Luo Y K.The no-structural carbohydrate of stem and sheathin relation to panicle characteristics in rice[J]. Chinese Journal ofRice Science ,2001,15(2):155 157.