
下载掌阅APP,畅读海量书库
立即打开



上面讲了我们的感觉器官主要只对在时间上有变化的刺激才有反应,那么对空间上的变化是不是也是这样呢?美国视觉科学家哈特兰(Haldan Keffer Hartline)(图 2.13)通过几十年不懈的研究,证明了在生物的感觉系统中相近的神经细胞之间存在相互抑制作用,他把这种来自侧向的抑制称为侧抑制,正是这种侧抑制作用加强了空间中有变化的部分。

图 2.13 哈特兰
哈特兰的父亲是位生物学教师,年轻时哈特兰常和父亲在大西洋岸边漫步,父亲不时会指给他看一些奇奇怪怪的生物,其中一种给他留下了特别深刻的印象。这是一种甲壳动物,叫做鲎。鲎的成体大约有一个脸盆那么大,后面拖着一条坚硬的尾巴[图2.14(a)],据说古时印第安人常常把它绑在棍子上当作长矛。在鲎的背部有两个腰圆形的复眼,每个复眼都由上千个小眼组成。这是一种非常古老的动物,历经沧桑而得以幸存下来,自然有它的优点。小哈特兰当时绝没有想到这种其貌不扬的家伙有朝一日会帮他赢得诺贝尔奖。
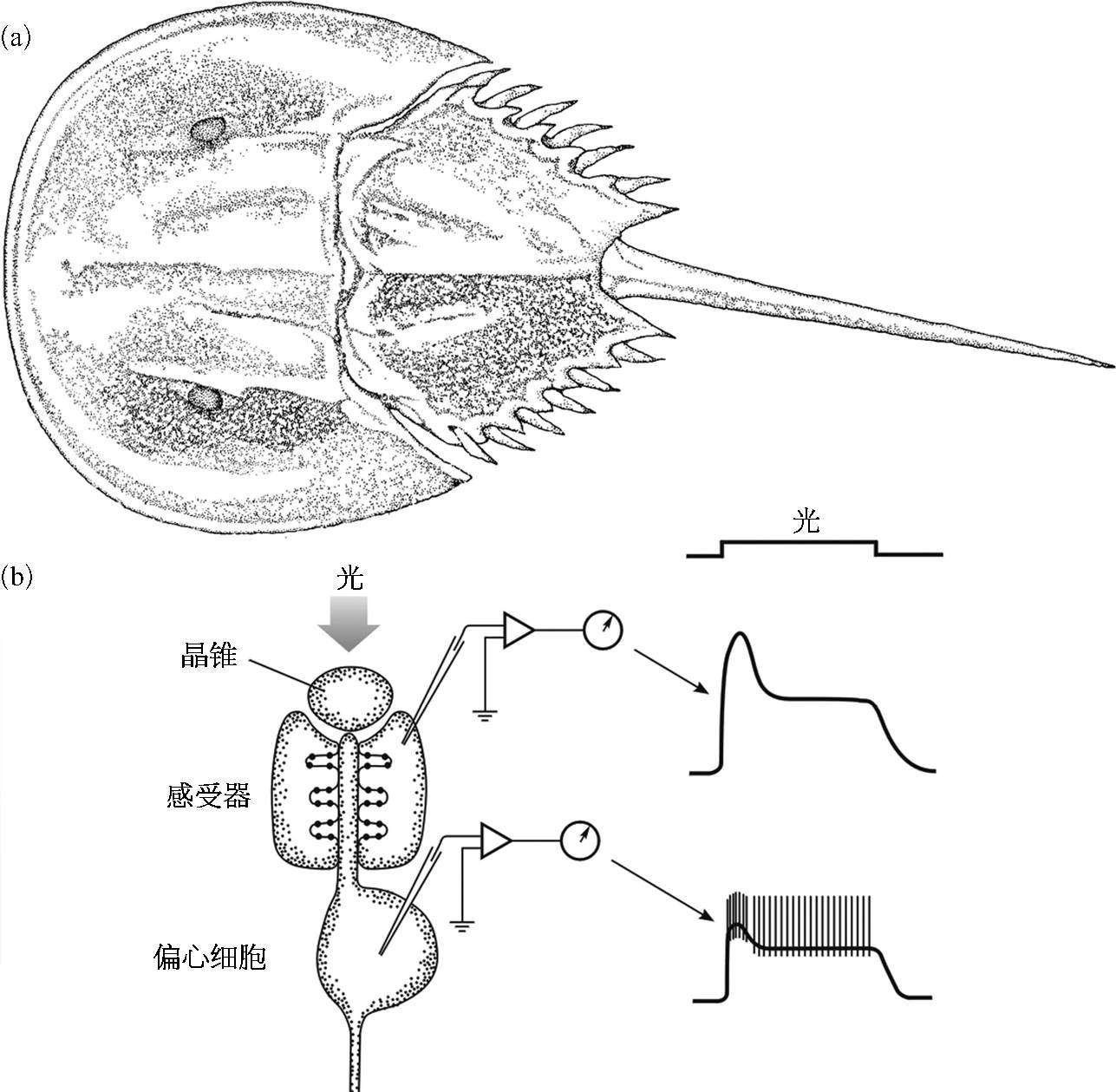
图 2.14 鲎 [5]
(a)鲎的外观;(b)鲎复眼上小眼的解剖结构,晶锥相当人眼中的晶状体。
哈特兰受父亲的影响选择学习生物学。他不但生物学实验做得很出色,对修理实验仪器也很精通。弦线电流计的弦断了,他就自己换!哈特兰对新技术和科学上的新潮流都很敏感。20 世纪 20 年代末,电子管放大器和示波器刚刚引入到生理学的研究,他就自己动手做了一台放大器,尽管外表不太好看,但用起来还是蛮不错的。
1927 年,艾德里安(Edgar Douglas Adrian,1932 年获得诺贝尔生理学或医学奖)在鳗的视神经上记录到了成群的神经脉冲,这引起哈特兰极大的兴趣。他想起鲎的光感受器很大,觉得它应该是研究单个神经细胞电活动的好材料。1931 年夏天,他前往伍兹霍尔的海洋生物实验室进行短期交流,专门研究鲎视神经上的电活动。一开始,他用幼鲎做实验材料,因为幼鲎经常要蜕壳,角膜显得非常清澈,而且其视神经也很容易分离。但是记录的结果总是不理想,一群脉冲杂乱无章,只是偶尔可以从中勉强辨别出单根神经纤维的活动,他很苦恼,但还是一个接着一个地做下去。就在他临走前两天,所有的幼鲎统统用光了,只剩下两只成鲎,只好将就着用了。这两个家伙个子很大,背壳粗糙,两眼呆滞,而且擦伤很严重,怎么看都不讨人欢喜。谁知道真是“鲎不可貌相”,它们的视神经不仅很容易分离,而且当光照到它们的复眼上时,在视神经上可记录到一串很有规则,而且峰值很高的脉冲[图 2.14(b)]。一个夏天辛勤的劳动终于得到了丰硕的回报!他发现不管光照的强度如何,所记录到的神经脉冲的形状和幅度都一样,不一样的是单位时间里神经脉冲的个数(发放率)。换句话说,光照强度是用神经脉冲的发放率来编码的。以后,他又发现这种发放率与光照强度的对数成正比,这和我们人主观感觉到的亮度变化是一致的。
有一次,哈特兰做实验的时候,发现一根视神经的脉冲发放突然变稀疏了,原来有光漏到了其他小眼上。这种现象他以前也常看到,但他通常只是把这作为一种干扰,设法不漏光就是了。可是这次,他突然灵光一闪:“这不正说明各个小眼的活动不是彼此独立的,而是一个小眼的活动会影响另一个小眼的活动吗?”也就是说,一个小眼的活动会抑制其相近小眼的活动。想到这里他非常兴奋,他记起以前看过的鲎小眼层后面的神经纤维丛的染色照片。鲎的视网膜一共只有两层细胞:感光细胞和发放脉冲的偏心细胞。偏心细胞的轴突就是视神经[图 2.14(b)],视神经在传向脑的过程中,发出许许多多的侧枝,这些侧枝与其他小眼的视神经形成突触联系,这种联系必定是小眼之间相互影响的结构基础!哈特兰切断这些侧枝,果不其然,当光照邻近小眼时,他所检测的那个小眼的脉冲发放不再减少,也就是说,邻近小眼对它的抑制消失了。
哈特兰意识到幸运之神正在向他招手,他正处于一个重大发现的前夜。于是他把研究方向从研究单个细胞的活动转到研究神经细胞之间的相互作用。开始研究一种新的现象时,最好把情况尽量简化,所以哈特兰最初只研究两个小眼的情况。他先在第一个小眼上加上固定强度的光刺激,让第二个小眼完全对光屏蔽,测量其神经纤维上的脉冲发放率;然后在第二个小眼上加上不同强度的光刺激,再测量第一个小眼的脉冲发放率,这两者的差值就代表第一个小眼被第二个小眼抑制的程度。他用坐标图记录下这个差值与第二个小眼所受到的光照强度之间的关系,想从中发现规律,但结果很不理想。后来他想到,在第一个小眼受到第二个小眼的抑制时,第二个小眼同时也要受到第一个小眼的抑制,这种抑制作用是相互的。因此,应该考察受到抑制的量和施加抑制的小眼当时的活动程度(而不仅仅是它当时所受刺激的强度),也就是说它的脉冲发放率之间的关系。搞清楚了这一点,哈特兰很快就把这种关系的数学方程式写出来了。以后,他又把这种关系推广到多个小眼之间相互作用的情况。他把实际测量得到的结果和根据方程式计算得出的结果进行比较,两者符合得非常好[图2.15(b)]。由图 2.15 可看到,尽管加在鲎复眼上的光刺激是相邻的两块明暗不同的均匀方条,但是鲎复眼视神经上脉冲发放率的分布却不这样,在明暗交界处的两侧分别出现峰和谷,这正好和我们人的一种错觉——马赫带现象是一致的,即在明暗交界处的两侧会分别“看到”一条特别亮的亮线和一条特别暗的暗线(关于这点,笔者在下一章中还要详细讲)。
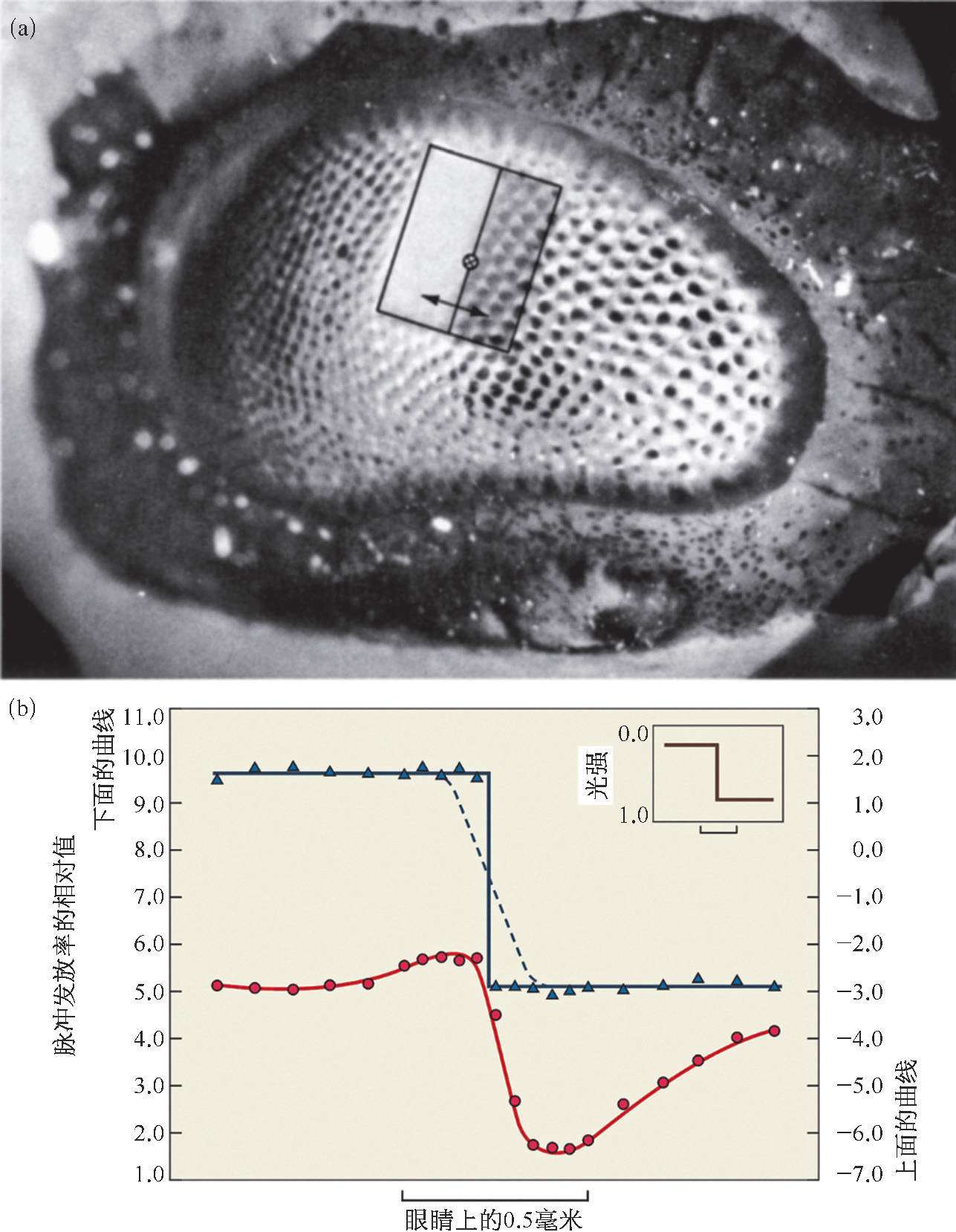
图 2.15 鲎的小眼之间的相互作用实验
(a)在鲎复眼上加上有明暗对比的光刺激。(b)右上角的折线表示光刺激的光强变化,圆点表示这种光刺激所引起的各个小眼的脉冲发放率的实际测量值,圆点所在的曲线表示根据方程式计算得到的结果。如果每次仅暴露一个小眼,而把所有其余小眼都遮蔽起来,也就是说小眼没有受到附近其他小眼的侧抑制,在同样光刺激下测得的结果用三角表示,三角形所在的折线是计算所得的结果。虚线表示由于光感受器前面的曲光体(晶锥)对光线造成弥散,所以明暗分明的光线照射到复眼时,实际落在光感受器上的光分布在分界处形成一个过渡区,这就解释了为什么圆点和曲线在分界处不那么陡。
哈特兰的研究结果不仅明确地显示出在生物的神经系统中确实存在着各个组成细胞之间的相互抑制作用,而且这种网络还会表现出类似马赫带的现象,这就给这种心理错觉提供了坚实的神经生物学基础。哈特兰解释说,这种相互抑制作用的功能很可能就是把图像的边缘突显出来,使生物易于检测空间中有变化的部位。这些部位正是携带着关键信息的部位。例如,一张非常传神的漫画寥寥几笔就能使我们认出画的是谁,其效果并不比看到这个人的照片差多少(图 2.26)。
事实上,只有有变化的地方才有信息,没有变化也就没有信息。所以侧抑制很可能就是把这种有变化的地方突显出来,使神经系统在传送信息时更为有效。哈特兰由于上述一系列研究而荣获 1967 年的诺贝尔生理学或医学奖。他和霍奇金一样,也在获奖感言中强调动物标本的选取十分重要,鲎在他的发现中作出了巨大贡献。他说:“我们选择鲎作为材料真是一大幸事。”“这个视网膜刚刚复杂到足以引起人们的兴趣,然而看起来又简单到足以使人们相信最终是可以把它搞清楚的。”善于选取合适的标本也是科学研究的一大艺术吧!
以后的研究还发现,不仅像鲎这样原始的动物的视觉系统中有侧抑制作用,而且包括人在内的高等动物的各种感觉系统里面都有这样的作用。现在就拿其他动物的视网膜作为例子来说明这个问题。图2.16 是拉蒙— 卡哈尔笔下的视网膜的解剖结构。这张图虽然已经有 100年左右的历史,但是直到今天大致上还是正确的。可以看到这种视网膜至少有三层结构,而不是像鲎的视网膜那样只有两层结构。一个神经节细胞可以接受一个或者多个双极细胞的输出,而一个双极细胞又可以接受一个或者多个感光细胞的输出。在双极细胞和神经节细胞之间有无长突细胞横向联系;在双极细胞和感光细胞之间则有水平细胞在横向联系。所以对某个神经节细胞来说,刺激和神经节细胞有联系的不同部位的感光细胞都有可能改变它的反应。视网膜上受到刺激能对某个神经节细胞的活动产生影响的这种区域就称为这个神经节细胞的感受野。后来的研究发现哺乳动物神经节细胞的感受野一般呈同心圆结构。刺激感受野的中心部位和刺激感受野的周边部分,其作用正好是互相拮抗或者说互相抑制的,因此这里就有某种侧抑制作用。一般认为,这种作用有利于视觉系统提取图像的边框,因此在形状知觉中起重要的作用。
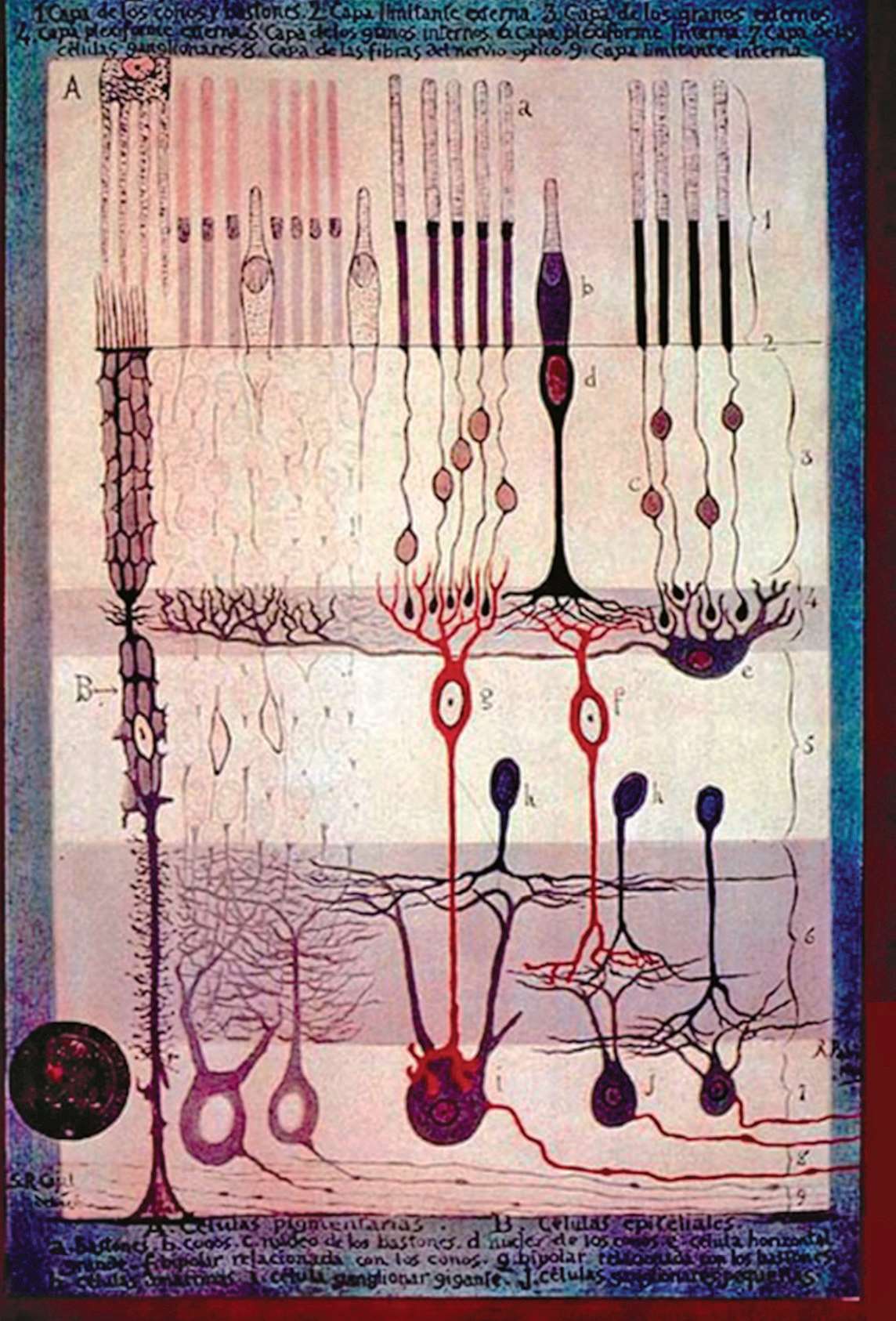
图 2.16 拉蒙— 卡哈尔笔下的视网膜解剖图
a:视杆细胞;b:视锥细胞;g、f:双极细胞;e:水平细胞;h:无长突细胞;i、j:神经节细胞。神经节细胞的轴突(图片最底下的一些水平走向的细线)就构成了视神经。进入眼中的光线是从图底下往上传的,所以光线要先穿过视神经和一系列神经细胞,最后到达感光细胞。