
下载掌阅APP,畅读海量书库
立即打开



(1)湖泊极端水文事件的定义与识别方法
湖泊极端水文事件指在特定时间尺度上湖泊水文过程发生的小概率事件,一般具有相对水文极值、持续一定的时间、对湖泊水安全和水生态环境产生了严重的影响等特征(Beniston et al.,2007;Lei et al.,2019;张利平等,2011)。研究湖泊极端水文事件最常用的水文变量是湖泊降水和水位,这些变量相对易于获取且时间序列较长,对湖泊水文特征的描述最为直观(Alsdorf et al.,2000;Carter et al.,2018;Li et al.,2018;Zhang et al.,2018;李云良等,2015;王鹏等,2014)。随着遥感技术和水文水动力模型的不断发展,湖泊水面积、蓄水量、流速、洲滩湿地的淹没面积等水文变量也逐渐被用来研究湖泊极端水文事件,这些变量的使用对于进一步探索极端水文事件对湿地生态水文过程、湖泊水生态系统和水环境的影响具有非常重要的意义(Batt et al.,2017;Buma et al.,2019;Li et al.,2019;Tan et al.,2019;Zhang G et al.,2017;边多等,2006)。
目前,关于湖泊极端水文事件的识别并没有统一的标准,其中湖泊水文极值的确定是湖泊极端水文事件识别的关键内容。湖泊水文极值的确定方法主要包括三种:①经验法,主要根据湖泊水文变量(通常指水位)大小对当地生产和生活产生的影响来确定(Bolgov et al.,2017;Li et al.,2017;Yao et al.,2016;闵骞等,2010);②极值法,特定时间段内湖泊水文变量的最大/最小值(Carter et al.,2018;Klamt et al.,2020;Lei et al.,2019;Shankman et al.,2006;Zhang et al.,2018;Zhang et al.,2019;Zhu et al.,2019);③极值分布函数,水文变量的分布超过一定的概率,如广义极值分布函数、皮尔逊Ⅲ型函数、韦布函数等(Jalbert et al.,2019;Liu et al.,2016;Paynter et al.,2011;Zhang D et al.,2017)。根据研究的区域和时间尺度,不同的学者选择不同的湖泊水文变量和水文极值识别方法来识别相应的湖泊极端水文事件,进而对湖泊极端水文事件的基本特征进行分析,主要包括:极端事件的起止时间、持续时间、严重程度、峰值、发生频率、幅度、变化趋势等,这是湖泊极端水文研究中讨论最为广泛的部分(Buma et al.,2019;Kienzler et al.,2015;Lei et al.,2019;Liu et al.,2016;Nandintsetseg et al.,2007;Paynter et al.,2011;Shankman et al.,2006;Sun et al.,2018;Zhang D et al.,2017;Zola et al.,2006;孙占东等,2015;姚静等,2017;赵化雄,2003)。
(2)湖泊极端水文事件成因的识别
湖泊极端水文事件的产生是流域入/出湖水文过程和湖泊自身水量收支过程共同作用的结果(Carter et al.,2018;Ghale et al.,2018;Lei et al.,2019;Satge et al.,2017;Sun et al.,2013;Zhang et al.,2014)。流域水文过程受到气候、土地利用/覆被变化和社会经济发展用水的综合影响,湖泊本身作为陆—气交互作用的特殊界面,水热条件(降水和蒸发)剧烈波动,加之湖区人类活动的影响,使得湖泊极端水文事件的成因异常复杂。常用的湖泊极端事件成因的识别方法可分为统计分析法和数值模拟法,其中统计分析法主要用于湖泊极端水文事件成因的定性识别,包括相关分析法和联合分布函数法(Assani et al.,2016;Biron et al.,2014;Ghale et al.,2018;Tian et al.,2014;Zhang D et al.,2017;郭华等,2012;李景保等,2011);数值模拟法主要用于湖泊极端水文事件成因的定量识别,包括机器学习法和水文水动力方法(Liu et al.,2018;Myronidis et al.,2012;Shiri et al.,2016;Sun et al.,2015;Zola et al.,2006;李云良等,2015;姚静等,2017)(图1.1)。此外,基准期(未发生极端事件的时期)的选定是研究湖泊极端水文事件成因的前提,它直接影响到极端水文事件的归因结果。基准期的选定有两种:一是选择没有发生湖泊极端水文事件的历史时期;二是模拟没有发生湖泊极端水文事件时的假定情景。
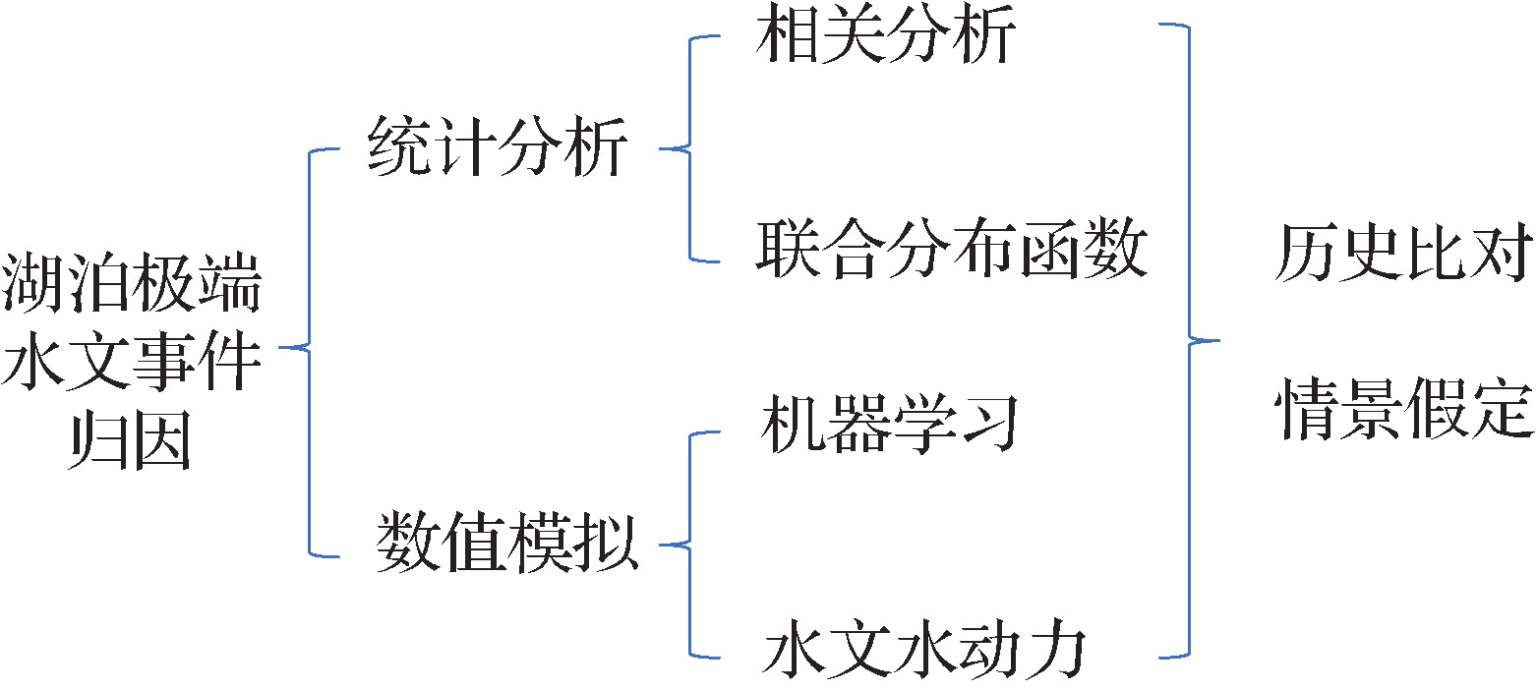
图1.1 湖泊极端水文事件成因识别方法
大量的学者采用上述方法研究了流域来水、湖泊降水和蒸发、湖泊出流对极端水文事件的影响。其中,流域来水和湖泊本身的水量收支(降水和蒸发)作为极端水文事件产生的水分来源,是影响湖泊极端水文事件的关键因素,它们均受气象条件和人类活动的影响。为了更好地探讨湖泊极端水文事件的成因,粗略地将湖泊极端水文事件分为湖泊洪水事件和湖泊干旱事件。就湖泊洪水事件而言,其发生的主导因素一般是气象条件,即发生了极端来水或湖泊本身发生了极端降水事件(Abbasi et al.,2019;Bing et al.,2018;Carter et al.,2018;Li et al.,2016;Riboust et al.,2015;Shankman et al.,2012;匡燕鹉等,2019;叶正伟,2006)。比如,Riboust等(2015)采用水文水动力模型,发现气象条件是影响2011年安大略湖洪水事件的主要因素;匡燕鹉等(2019)采用统计分析法,发现2017年洞庭湖特大洪水是由流域极端来水和湖区极端降水共同引起的。相较于湖泊洪水事件,湖泊干旱事件的成因比较复杂。这是因为湖泊干旱事件涉及的各因素之间有着强烈的陆—气交互作用,且干旱事件的持续时间较长,影响范围较广,加之湖泊类型、所处气候区和研究尺度的差异,不同的研究得出的湖泊干旱事件的主导因素有所差异。主要包括:①气象条件主导型。气象条件(降水、冰雪融水)引起的流域来水减少、湖泊降水减少和蒸发增加,是湖泊干旱发生的触发器,这种类型的湖泊干旱事件可发生于全球所有气候类型的湖泊(Tian et al.,2014;Yao et al.,2016;Zhang et al.,2009;刘元波等,2014)。比如,Zhang等(2009)采用统计方法,发现美国中部平原地区小型湖泊的干旱事件主要受到冰雪融水引起的入湖径流减少和蒸发增加的影响;Yao等(2016)采用水文水动力模型,发现流域降水减少引起的入湖径流降低是1963年鄱阳湖春季干旱发生的主要原因。②人类活动主导型。人类活动一方面影响入湖径流的大小(下垫面改变、人类取用水和水库调蓄),另一方面直接影响湖泊的蓄水量(湖区人类取用水和围垦),这种类型的湖泊干旱事件主要发生在水资源比较匮乏、人类活动强度较大的干旱区湖泊(Ghale et al.,2018;Satge et al.,2017;王青等,2013)。比如王青等(2013)通过统计分析法,发现人类活动引起的入湖径流量减少是白洋淀发生干旱的主要原因;Ghale等(2018)采用统计分析法,发现人类活动对1999—2010年伊朗西部的乌尔米亚湖干旱事件的贡献量为72%~87%。值得注意的是,对于过水型湖泊和吞吐型湖泊而言,除了入湖水文过程、湖泊降水和蒸发的影响外,湖泊出流过程也是影响极端水文事件的重要因素(Biron et al.,2014;Li et al.,2017;Li et al.,2016;Yao et al.,2016;赖锡军等,2008)。以鄱阳湖为例,Yao等(2016)发现2006年秋季的鄱阳湖干旱是因为长江来水减少引起的拉空作用使得湖泊出流增加。
(3)未来发展
总的来说,湖泊极端水文事件的研究取得了丰富的成果,这对区域水资源管理与湖泊水灾害事件的应对提供了科学参考。在以后湖泊极端水文事件的研究中,可以加强开展以下几个方面的工作:
①自然波动和人为因素对入湖水文过程的影响方式与强度。入湖水文过程直接影响湖泊水文过程。气候变化、土地利用/覆被变化、社会经济用水和水库调蓄作为影响入湖水文过程的外部驱动力,均受到自然波动和人为因素的双重胁迫,尤其在人口众多、经济较为发达的湖泊—流域系统中,人为因素对入湖水文过程产生了深远的影响。辨识自然和人为因素对入湖水文过程的影响方式与程度,是从“山水林田湖草”出发,科学管理和调控湖泊—流域系统水安全的重要内容。
②外部驱动力、流域水文极值与湖泊水文极值之间的传递与响应关系。对湖泊—流域系统而言,外部驱动、流域水文过程、湖泊水文过程具有显著的联动效应,气候、植被、社会经济用水、水库调蓄多因素共同作用于湖泊—流域系统的关键水文过程,引起湖泊水文过程在较短的时间尺度上发生剧烈波动。因此,明确外部驱动力、流域水文与湖泊水文之间的传递与响应关系,是揭示湖泊水文极值生消机理的关键环节。
水文条件变化是控制湿地特征的关键因子,是湿地典型的扰动特征之一(徐治国等,2006)。湿地水文条件对湿地生态系统结构和功能具有重要的影响作用,直接控制着湿地生态系统的形成和演化(邓伟等,2003),控制着许多生态学格局和生态过程,特别是控制了基本的植被分布格局(黄奕龙,2003;李旭等,2009),是湿地生态系统演替的主要驱动力(Maltchik et al.,2007)。湿地水文条件的轻微改变,如湿地水分条件和水位变化将直接影响湿地物种的萌发、存活、生长和繁殖(Dwire et al.,2006;章光新等,2008;陈敏建等,2008;Hammersmark et al.,2009)。研究发现,土壤近饱和或地表浅积水(0~3 cm)是芦苇根茎萌发的最适宜环境条件,地表水深0 cm时芦苇的萌发率达90%,淹水4 cm时萌发率降低至60%,而淹水15 cm时仅2%(Yu et al.,2012)。此外,淹水时间和频率也会影响物种的萌发和幼苗生长。Casanova等(2000)研究发现,完全不淹水条件下幼苗的生物量最大,而始终淹水显著降低了幼苗的植株密度和生物量,短频率的淹水会促进萌发期的物种丰富度。
水文条件变化也会影响湿地植物的生理活动,如光合作用、呼吸速率、细胞内的叶绿素和酶含量等。Li等(2004)研究发现,淹水和间歇性淹水有利于提高香蒲的净光合速率、气孔导度和生物量,间接性干旱会降低香蒲叶片叶绿素含量和生物量。Pagter等(2005)研究了不同水分胁迫程度对芦苇生理生态特征的影响,结果显示,极端干旱胁迫下芦苇CO 2 同化速率和蒸腾速率显著减小,游离脯氨酸和渗透压增大,最终导致叶面积指数大大减小。Vretare等(2001)对芦苇和莎草等湿生植物的研究表明,地上地下生物量比值与根茎直径会随着水深的增加而显著增大,同时发现,深水环境生长的芦苇株高更高,根茎埋深更浅,Engloner(2004)也有类似的发现。
水文条件的改变还可以影响植被空间分布格局和演替过程(李胜男等,2008),导致生物区系在物种组成、物种丰富度和多样性以及生态系统生产力方面较大幅度的变化(Mitsch et al.,2000)。研究发现,湿地植物种具有沿水位梯度分布的格局,随着水位梯度的增加,植被群落优势种将经历“旱生—湿生—水生”的转变(田迅等,2004)。如Watt等(2007)对地中海地区的季节性洪泛湿地的地下水位进行了实时监测和植被调查,结果表明夏、秋季节的平均地下水埋深是决定植被组成和分布的主要环境因素。Bornman等(2008)通过典范对应分析发现南非河口湿地植被空间分布主要受土壤含水量和地下水埋深的共同影响。此外,周期性的高低水位交替变化有利于维持湿地生物多样性,有利于r策略物种的生存,这一结论已基本成为生态学界的共识(Riis et al.,2002)。进一步研究发现,流水环境及周期性高低水位交替变化可显著提高湿地的初级生产力,有利于维持湿地的生物多样性(Wagner et al.,2000)。而水位波动幅度和持续周期不同时,水位波动的影响效果也存在明显差别。如Riis等(2002)对新西兰21个湖泊湖滨植被群落的调查发现,当水位变幅接近1 m时,30 d左右的出露期对维持当地植物群落物种多样性最为有利。Blanch等(1999)的研究发现,湿地植物丰富度与水位波动过程中水深处于0~60 cm的淹没时间及1 m以上的出露时间相关。
而极端的水文条件变化,如长期洪水或干旱,易使湿地生态系统发生质的变化,向水生或陆生生态系统转变,同时伴随植物物种的减少和生物多样性的下降(Bond et al.,2008)。如1998年大洪水导致鄱阳湖区水生植物生物量骤降,物种减少,湖区初级生产力降低。Gacia等(1996)发现,在一次持续时间较长的洪水过后,原水韭属植物的生长位置上移,新的生存高度相应提升。Vandervalk等(1994)对水位永久上升后湿地植被的调查表明,样区的物种数、根系密度和Shannon-Wiener多样性指数全面下降。而频发的干旱及其滞后作用也会对湿地植被的生长、演替以及生态系统结构和功能属性造成严重影响(Berdugo et al.,2020)。干旱期间水文连通性降低,阻碍了湿地斑块间的物质循环和能量流动,湿地景观类型会依据水文梯度发生变化(余新晓等,2006)。随着干旱的持续,一些植物失去适宜生态位,造成湿地植被大面积死亡,植物群落构成发生变化,稳定性降低,原有湿地景观萎缩,并向均质化和旱化的方向发展(章光新等,2008;陈敏建等,2008)。持续性严重干旱甚至会直接改变整个湿地生态系统的原有结构,导致湿地植被生产力的下降和生态系统碳的流失(Huang et al.,2016),进而导致生态系统各项功能下降(Griffin-Nolan et al.,2018)。例如,Laine等(1995)对排干3~55年的不同沼泽湿地进行研究,发现了以莎草种消失为显著特征的沼生植被向森林植被演替的过程,同时伴随着整个研究区的γ多样性显著降低。
总的来说,目前的研究对系统、全面地认识湿地植被对不同水文要素变化的响应产生了积极的作用。相对于周期性的水文要素波动变化而言,非周期性的极端水情变化对湿地生态系统的影响更大,会使湿地植被系统发生质的变化,但目前的研究涉及极端水情的不多,如何确定长时间淹水、干旱胁迫等极端水情条件对湿地植被时空分布格局和演替过程的影响以及如何确定典型湿地植被群落对于这些水情条件的耐受阈值,仍有待于进一步研究。
陆地生态系统的碳循环过程通过一系列生物地球化学过程,吸收或释放CO 2 和CH 4 等温室气体,控制大气以及地表的能量和水分交换,从而显著影响未来的气候状态,已成为当前全球变化研究的一个热点问题(朴世龙等,2019;张婷等,2022;曹明奎等,2004;Lu et al.,2018;He et al.,2019;杨元合等,2022;Krüger et al.,2022)。陆地生态系统的净初级生产力(NPP)指绿色植物在单位面积、单位时间内所累积的有机物数量,表现为光合作用固定的有机碳中扣除植物本身呼吸消耗的部分,这一部分用于植被的生长和生殖,也称净第一性生产力。NPP作为地表碳循环的重要组成部分,不仅直接反映了植被群落在自然环境条件下的生产能力,表征陆地生态系统的质量状况,而且是判定生态系统碳源/碳汇和调节生态过程的主要因子,在全球变化及碳平衡中扮演着重要的作用(方精云等,2001;Running et al.,2004;Xu et al.,2019)。
由于植被NPP在陆地碳循环中的重要性,所以准确估算植被NPP至关重要。早期的NPP估算主要基于实地站点测量,测量方法主要有直接收割法、光合作用测定法、微根管法、CO 2 测定法等(贺金生等,2004;Ni,2004)。然而,实测法虽精度较高,但是其只能适用于点式NPP测定及小范围观测。模型模拟法是通过构建数学模型来进行生态系统碳循环模拟和生产力定量估算的方法,方法机理性强但模型参数数据不易获取。随着对陆地生态系统NPP研究工作的深入开展,以及航天、遥感、GIS和计算机技术的快速发展,大量基于遥感的NPP估算模型先后出现,为区域和全球尺度的NPP估算及相关应用研究带来了新的活力。到目前为止,各种建立的碳循环模型大致可以总结为四类:气候生产力模型、生态过程模型、光能利用率模型以及生态遥感耦合模型(洪长桥等,2017)。现如今,国内外针对陆地植被生产力的研究已然取得了长足的进展,但是由于陆地表层系统的高度空间异质性,不同模型在参数、结构方面的差异以及输入数据的影响,碳循环模型的模拟结果往往存在较大的不确定性(Verbeeck et al.,2006;Cramer et al.,2010)。目前,相对成熟的光能利用率模型,由于区域尺度转化容易,所需参数较少,通过遥感可以直接获得大量输入数据,在大范围生态系统模拟中具有一定的优势和可靠性,是陆地生态系统生产力估算比较好的方法之一(赵国帅等,2011)。此类模型的代表有CASA、GLO-PEM、SDBM等,其中,CASA模型由于操作简单、精度高而被广泛应用于不同区域和尺度的NPP估算(董丹等,2011;孟元可等,2018)。
植被NPP受气候条件以及下垫面土地利用/覆盖变化、生态工程建设、城市化扩张等多种因素的影响,其动态变化过程相当复杂。气象因素决定了区域水热条件,直接影响植被生长。研究表明,在全球尺度上,陆地生态系统生产力呈纬向分布规律,低纬度地区高于高纬度地区,纬度每升高1°,NPP减少11.05 g C·m -2 ·a -1 (Ji et al.,2020;Wu et al.,2020)。主要气候因子中,温度与植被生长呈正相关,而降水的影响则随地区湿度而变化(Yu et al.,2013;Xu et al.,2014)。在干旱半干旱地区,植被生长主要受降水控制,温度和辐射的影响相对较弱(Austin et al.,2006)。在机制上,气候变化对植被生产力的影响主要通过影响光合作用、呼吸和土壤有机碳的分解过程来实现(Chiew et al.,1995;Schreider et al.,1996)。人类活动作用是引起植被NPP变化的另一个重要驱动因素。土地利用变化和城市化通过改变陆地生态系统的碳储量和通量进而影响碳循环(Hutyra et al.,2011)。城市化是人类活动引起植被NPP变化的关键驱动因素之一,其作用机制也很复杂。研究表明,1992年至2000年,由于城市化的发展,美国东南部的年均NPP下降了0.4%(Milesi et al.,2003)。在中国长江三角洲的城市地区,城市化进程在1999—2010年造成了4.7 g C·m -2 ·a -1 的NPP下降趋势(Wu et al.,2014)。
值得注意的是,20世纪中期以来,以全球变暖为主要特征的气候变化,使得全球和区域水分循环加速,导致极端气候干湿事件的频率和强度明显增加,对陆地生态系统的结构和功能产生了深远的影响,已成为当今研究的热点(朴世龙等,2019;Jiang et al.,2020;齐贵增等,2021)。Hilker等(2014)指出自2000年以来,降雨量的减少已经降低了亚马孙大部分地区的植被绿色度,在厄尔尼诺事件期间,NDVI在高达10%的范围内降低了约16.6%。John等(2013)对蒙古高原的研究发现,沙漠生物群落比草原生物群落更容易受到干旱的影响。Song等(2019)发现我国西南地区 2009—2010年冬春干旱期间植被覆盖度和NPP下降,2011年夏季干旱期间植被变绿,NPP增加。尽管不同陆地生态系统对极端气候的敏感性不同,但干旱在不同陆地生态系统中无疑是驱动NPP减少的主要因素之一。如果气候变化诱导的干旱持续增强,将会通过“碳—气候”正反馈调节而加速全球变暖(Ivits et al.,2014;Ma et al.,2012),从而产生更加严重的干旱,并且可能会导致生态系统不可逆的退化。干旱同时也影响生态系统中的植被生长,不同植被在应对干旱时的不同生理反应决定了对水亏损的抵抗力稳定性和恢复力稳定性的水平(Vicente-Serrano et al.,2013)。极端气候事件对生态系统具有较大的负面影响,彻底理解生态系统对极端气候变化的响应,不仅对提高全球变化情景下植被转移预测精度具有重要理论意义,而且对提高气候波动和气候变化对植被造成的脆弱性认知具有重要的科学价值。
湿地生态系统作为陆地与水体的过渡生态系统类型,与海洋、森林并称为全球三大生态系统,在世界各地分布广泛,约占全球陆地总面积的6%(刘红玉等,2003)。相比于其他生态系统,湿地生态系统具有更高的生产力和碳元素密度,全球湿地总碳储量约占陆地生态系统总储量的14%(Sharifi et al.,2013)。湿地的水文特征独特,既不同于排水良好的陆地生态系统,也相异于开放式的水生生态系统,其独特性也使得湿地系统在防御洪水、净化水质、涵养水源、固碳释氧、调节区域气候、保护物种多样性以及维护生态系统平衡等各个方面均发挥着不可替代的重要作用(崔保山等,2006)。目前,国内区域性NPP的模拟研究主要集中在青藏高原(沃笑等,2014)、内陆干旱区(龙慧灵等,2010)、西南(董丹等,2011)以及东北(Mao et al.,2014)等地的生态敏感区,植被类型主要集中于林地、草地、沼泽湿地等,这些重点研究区域植被NPP往往受气候变化(气温、降水、辐射等)影响较为显著。然而,与通常的陆地生态系统不同,水文条件的变化对通江湖泊洪泛湿地植被发育及NPP时空格局变化有着不可忽视的作用。当前,受制于高时空分辨率植被覆盖数据以及不同植被生物量获取的困难,有关长江中下游通江湖泊洪泛湿地植被NPP时空变化的系统研究还很少见。孟元可等(2018)曾利用多源遥感数据,并借助采用改进CASA(Carnegie-Ames Stanford-Approach)模型,模拟分析了2000—2015年鄱阳湖区NPP的时空变化状况。该研究指出,通过构建高时空分辨率卫星遥感数据驱动的生态模型,是研究高动态洪泛湖泊湿地生态系统NPP动态的最有效方法。
鄱阳湖洪泛湿地是典型的水陆界面频繁交换的湿地类型,湖泊水位变化是鄱阳湖湿地植被群落分布及演替的主控因子(胡振鹏等,2010)。湖泊水位的巨大变幅引起了鄱阳湖洲滩出露面积变化,进而决定了湿地面积在时间和空间上的剧烈变化(Feng et al.,2012;谢冬明等,2011),影响了湿地植被空间分布格局及生物量的变化(叶春等,2013)。同时,受水位梯度的影响,各典型植被群落占据特定的水分生态位空间,呈现出沿水岸线呈条带状、环状分布的总体格局(胡振鹏等,2010)。针对水情变化驱动鄱阳湖湿地植被生态响应,目前已开展了大量研究,如葛刚等(2011)通过野外调查发现,鄱阳湖洲滩湿地植被对水分梯度的依赖性是种群分布格局形成的主要原因。周云凯等(2017)利用实地测量数据,结合遥感与GIS技术研究了鄱阳湖湿地洲滩优势植物灰化苔草的固碳功能及其空间分异,发现灰化苔草全年固碳量约为88.15×10 4 t,年均固碳能力远高于其生长季内的碳释放能力,是鄱阳湖湿地的一个重要碳汇。他还进一步揭示了灰化苔草种群生产力对水淹条件和干旱胁迫的响应规律(周云凯等,2018)。张丽丽等(2012)以鄱阳湖自然保护区为研究对象,通过构建植被群落—水文参数直方图和计算敏感性指数,分析了不同植被群落对水文条件变化的耐受性和敏感性。余莉等(2011)、吴琴等(2012)、胡豆豆等(2013)、许秀丽等(2014)通过对鄱阳湖典型洲滩湿地不同植被类型地下水、土壤水的变化特征及生物量的测定分析,发现水位降低是导致苔草生物量减少的主要因素。王鑫等(2019)还研究了不同土壤湿度条件下鄱阳湖湿地3种优势植物芽库萌发和生长的影响,发现南荻和虉草芽的萌发率随土壤湿度的增加呈现先升高后降低的趋势,而灰化苔草芽的萌发率随土壤湿度的增加而显著下降。薛晨阳等(2022)对鄱阳湖湿地植物群落稳定性的最新研究发现,芦苇+南荻群落和狗牙根群落的稳定性与湖泊水情及洲滩土壤水分变化存在明显的关系。
除野外调查与原位监测研究外,部分学者还专门通过控制实验揭示湿地植被特征对特定水情变化的响应机制。如游海林等(2013)通过盆栽控制实验研究了鄱阳湖典型植被根系生长对极端水情条件的响应,结果显示极端旱化处理下植物主根长度显著高于须根长度。冯文娟等(2018;2020)开展了不同地下水埋深及淹水情景对灰化苔草种群生理生态指标影响的控制实验,发现株高和叶宽随着地下水埋深的增加而减小,而夏季淹水时长增加会阻碍秋季植被的生长恢复,但可能会对第二年春季的萌发产生促进作用。李文等(2018)、陈亚松(2020)也通过类似的控制实验分别研究了不同淹水深度、淹水时长对鄱阳湖洲滩湿地植物生长、功能性状及生物量的影响。
此外,在景观尺度,诸多学者还借助卫星遥感手段,通过对鄱阳湖湿地植被生物量和分布面积等进行遥感定量,从而揭示不同时期洲滩出露面积、植被空间分布与水位变化的响应关系。如叶春等(2014)利用长期卫星遥感数据,结合植被生物量野外调查,以2003和2006年极端干旱年份为例,从湿地植被面积、生物量密度和总生物量的角度分析了鄱阳湖湿地植被生物量对于极端干旱的响应。吴桂平等(2015)通过MODIS植被指数产品和同期的植被生物量调查,对鄱阳湖湿地2000—2011年植被生物量及生物量密度进行了有效重建,发现植被群落生长分布具有特定的季相变化特征,鄱阳湖水位的周期性涨落是影响其变化的一个重要扰动因子。张方方等(2011)、戴雪(2015)的研究也有类似结论。史林鹭等(2018)基于MODIS增强植被指数EVI数据,并利用EVI时间序列模型研究了水文连通性对鄱阳湖湿地植被覆盖和生产力时空动态的影响。杜飞(2018)基于Landsat TM/ETM + 遥感影像数据,还从鄱阳湖湿地生态景观变化的角度分析了其对低枯水位的响应特征。
目前,对于鄱阳湖湿地植被的研究成果颇丰,也为进一步揭示鄱阳湖湿地植被群落分布格局及其演替过程等提供了很好的基础。但总体来看,已有成果主要侧重于鄱阳湖湿地植被群落分布、景观格局及生物多样性等方面的研究,对洪泛湿地景观类型的转移过程、植被群落稳定性变化以及湿地植被NPP等研究存在不足,尤其是近年来鄱阳湖极端水情频繁出现,如何定量识别以洪涝、干旱等为主的极端水文情势对鄱阳湖湿地植被群落结构、功能及稳定性的影响是当前亟待解决的科学问题,具有现实紧迫性。