
下载掌阅APP,畅读海量书库
立即打开



线粒体生物合成的调控是一个复杂的生物学过程。核基因编码线粒体内与mtDNA复制、转录和翻译相关的所有蛋白质和酶类 [12] 。研究证实,大量核编码的蛋白因子参与了线粒体生物合成,包括NRF-1和NRF-2、过氧化物酶增殖激活受体 α (Peroxisome Proliferators-Activated Receptor α ,PPAR- α )和过氧化物酶增殖激活受体γ(Peroxisome Proliferators-Activated Receptorγ,PPARγ)、特异性蛋白-1(Sp1)和早期快反应基因产物、C-jun和C-fos。核呼吸因子 1和 2是转录调节子,作用于编码氧化磷酸化系统亚基组分的核基因的表达。同时,在核内它们通过绑定氧化磷酸化基因启动子的共有序列调节其他与mtDNA复制有关的基因的表达 [13-15] 。线粒体转录因子A(mtTFA)是一个转录因子,与mtDNA D-Loop区的启动子作用,调节线粒体基因组的复制和转录 [16] 。研究证实,mtTFA含有NRF-1和NRF-2一致的结合位点,从而为细胞整合核编码的蛋白表达和mtDNA编码的基因转录提供了独特的机理。越来越多的证据表明过氧化物酶体增殖激活受体γ共激活子 α (PGC-1 α )是一个重要的线粒体生物合成调节子 [17] 。在与转录因子网状系统的协同作用中,PGC-1 α 有助于协调有氧代谢相关基因的表达。很多核基因编码的线粒体酶类对PGC-1 α 产生应答,如NRF-1和NRF-2,他们触发编码呼吸链多肽及mtDNA转录和复制蛋白的核基因的表达。研究表明PGC-1 α 在适应性产热 [18] 和骨骼肌再生过程中 [19] ,其mRNA和蛋白水平的增加伴随着线粒体的增殖。转基因小鼠心脏、脂肪细胞和成肌细胞中PGC-1 α 的过表达可以导致线粒体的增殖。这些研究结果提示PGC-1 α 在调控线粒体生物合成和维持mtDNA过程中发挥重要作用,并且可能在外界刺激和线粒体生物合成间起着连接作用。
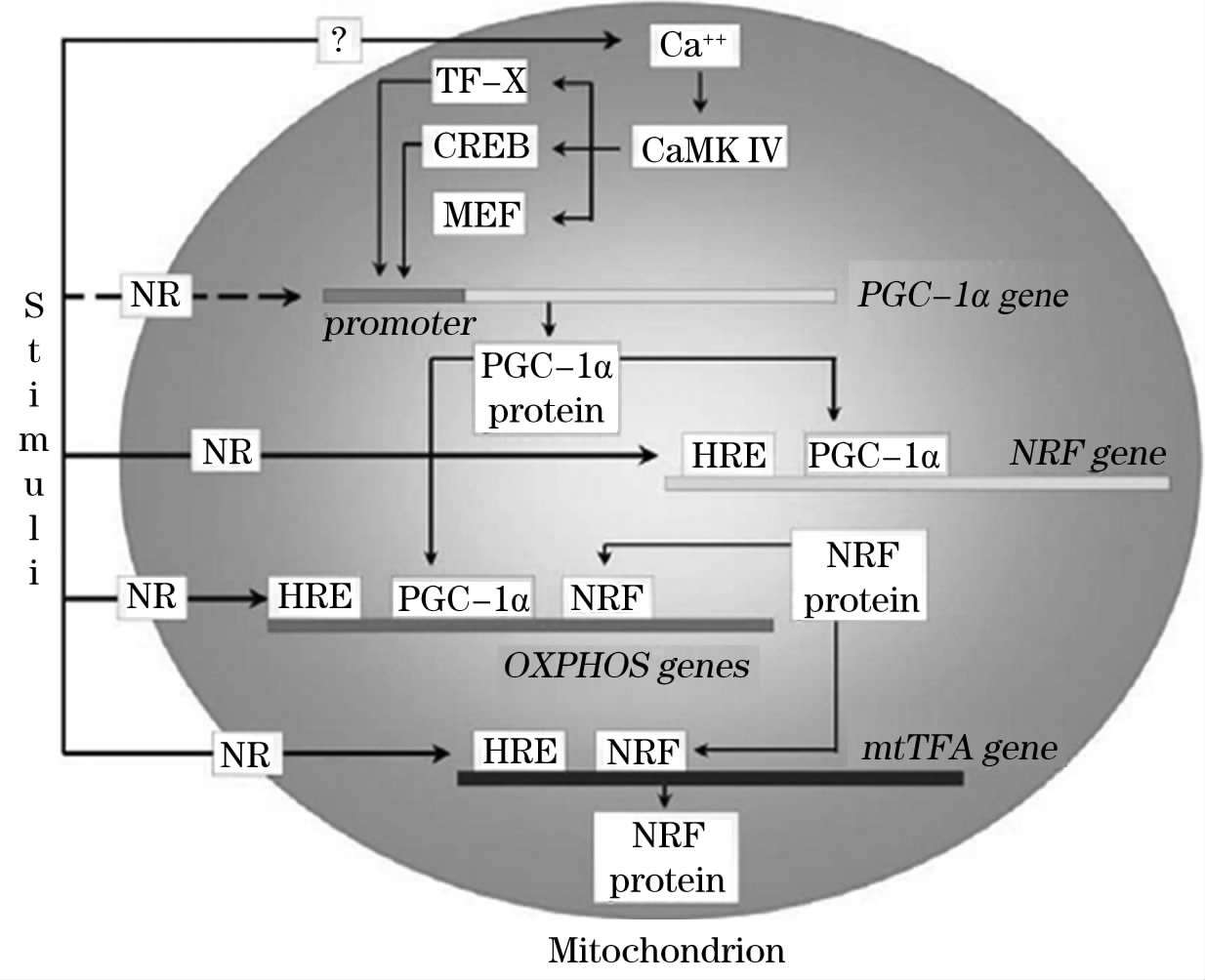
图1-1 线粒体转录因子调控线粒体生物合成 [20]
PGC-1 α 是近年来倍受关注的辅激活因子。1998年Puigserver等利用酵母双杂交技术从小鼠的棕色脂肪组织中发现了能与PPAR-γ相互作用的核辅激活因子PGC-1。小鼠暴露于寒冷环境中时,棕色脂肪组织和骨骼肌中PGC-1转录上调,其结合的转录因子PPARγ、甲状腺激素受体的活性增加 [21] 。随后,人和大鼠的PGC-1基因于 1999年和 2000年被相继克隆成功。PGC-1家族有 3个成员,PGC-1 α 以及它的两个共激活剂PGC-1β和PGC-1相关共激活剂PRC (PGC-1! Related Coactivator)。PGC-1 α 和PGC-1β优先在高氧化能力的组织,诸如心脏、慢收缩骨骼肌和棕色脂肪组织中表达,进而在线粒体功能能力和细胞能量代谢的调节中发挥关键作用。而有关PRC的表达模式和生物学作用还知之甚少。PGC-1 α 分子上具有几个重要的功能区域,其与一些特异靶器官的转录因子诸如NRs,NRF-1和MEF-2以及FOXO1相联系,同样还包含MAPK磷酸化位点 [22] 。
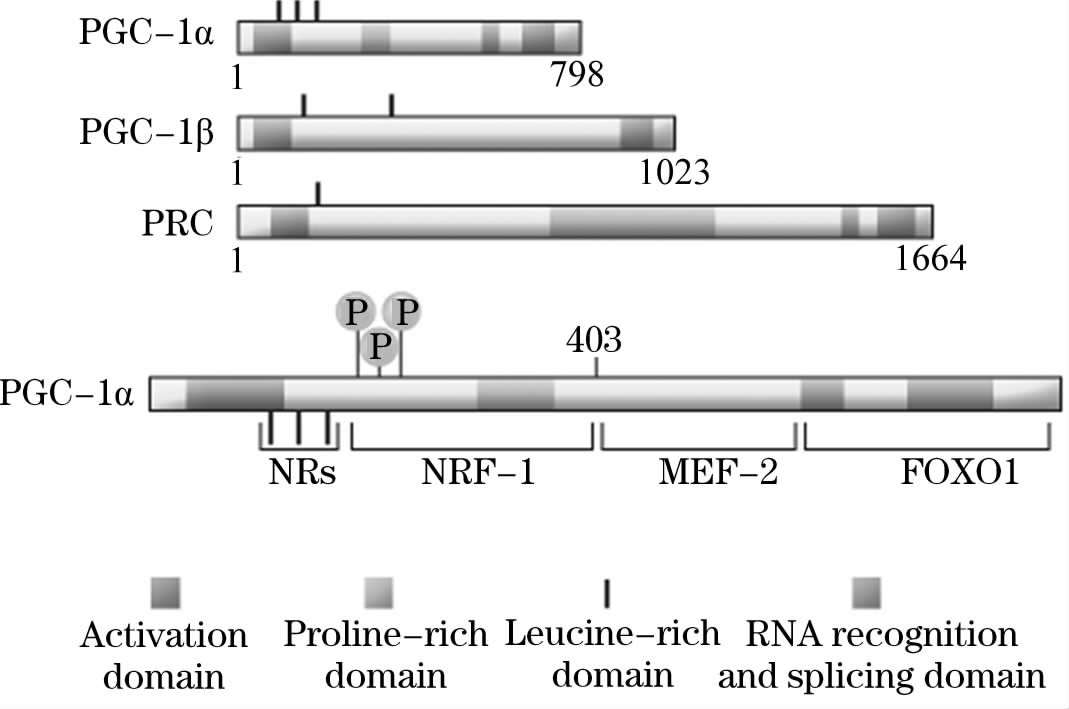
图1-2 PGC-1家族及其机构 [22]
PGC-1在斑马鱼、爪蟾以及哺乳动物诸物种间均具有相对保守的结构特征:蛋白的N末端含有一个转录激活域,其中含有主要的LXXLL基序(L为亮氨酸,X为其他氨基酸),也称为核受体盒(Nuclear Receptors Boxes,NR boxes),该基序有与具有介导配体依赖的核激素受体相互作用的功能;C末端含有一个RNA加工结构域包括RNA识别基序(RNA-Binding Motif,RMM)和富含丝氨酸/精氨酸的SR结构域(Serine-Arginine-Rich Domain),PGC-1氨基酸序列中还存在一个双向的核定位信号(Nuclear Bipartite Consensus Sequence,NBCS)。此外,在转录激活域和RNA加工结构域之间还存在一个抑制域,含有 3个保守的蛋白激酶A (PKA)作用的磷酸化位点,当体内cAMP浓度升高时,它们能被p38MAPK磷酸化,从而增强PGC-1介导的转录激活作用。PGC-1的结构组成提示其可能同配体依赖型或配体不依赖型的诸多核受体相互作用。此外,SR结构域和RNA识别基序共存于一个分子中,是PGC-1特征性的结构特点 [23] 。
PGC-1作为核转录辅激活因子,基于其核苷酸和氨基酸序列特点,通过多种分子机制发挥作用,从而多方面参与靶基因的转录起始调节,发挥其生物学功能。
PGC-1是哺乳动物能量代谢的关键调节因子之一,通过停靠许多转录因子,如NRF-1、MEF2C以及大多数核激素受体超家族成员,以不同的方式与其发生分子对接,进而调控其生物学功能。
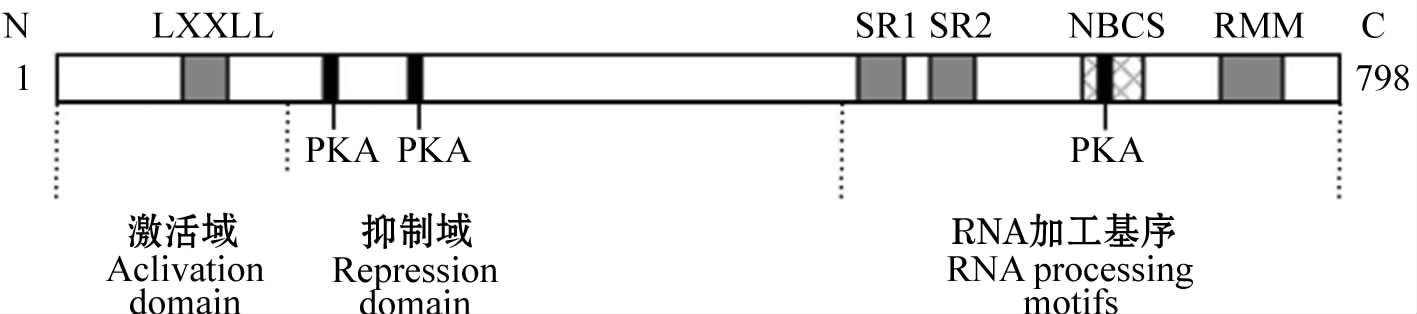
图1-3 人类PGC-1结构 [23]
注:LXXLL:富含亮氨酸的基序;PKA:蛋白激酶A磷酸化位点;SR:富含丝氨酸/精氨酸的结构域;NBCS:双向核定位信号;RMM:RNA识别基序。
辅激活因子停靠核受体超家族转录因子时,分子间相互作用存在配体依赖型和非配体依赖型两种模式。辅激活因子PGC-1以不同的分子作用模式停靠下游转录因子,从而发挥其生物调节作用。配体依赖型的作用模式中,PGC-1利用其LXXLL基序同核受体的转录激活功能域(Transcriptional Activation Function 2,AF-2)进行分子间的相互作用,这些核受体包括视黄醛酸受体(RXR α )、雌激素受体(ER)、甲状腺素受体(TR)、PPAR α 、糖皮质激素受体(GR)等。非配体依赖型的作用模式中,PGC-1停靠核受体完成转录激活作用不依赖相应配体的结合。肝核因子 4 α (HNF4 α )无需配体也能与PGC-1的LXXLL基序互作,表明即使没有外源性的配体,此核激素受体也处于激活的状态 [24] 。PGC-1也能利用不同的非LXXLL结构域同部分转录因子互相作用。PGC-1的第 200位至第 400位氨基酸间,存在与PPARγ和NRF-1相互作用的结构域;第 400位至第 500位氨基酸间,存在与MEF2C相互作用的结构域 [25] 。
PGC-1能与下游转录因子结合,从而促进靶基因转录进行。但是,转录因子启动的下游靶基因的启动子的特异性也决定PGC-1是否能与转录因子结合。辅激活因子不仅能增加转录效率,还在限定转录因子的特异性靶基因的环节中发挥重要作用。
核小体存在与否是基因表达的一个因素,核小体覆盖蛋白结合位点时,基因被关闭;核小体重新定位后离开该位点时,基因被打开。核小体的定位甚至可能是通过调控RNA聚合酶的进入而精细调控基因表达的关键因素。组蛋白乙酰化是改变核小体相对位置的方式之一。只有当核受体、辅激活因子和基础转录因子共同形成寡聚体复合物时,才能保证各种相互作用因子的稳定性和转录的特异性。大多数辅激活因子增加靶基因的转录活性是通过组蛋白乙酰转移酶活性使得染色质构象发生改变从而转录起始。但是,PGC-1不具有其他辅激活因子特征性的氨基酸序列,也不包含可识别的组蛋白乙酰转移酶(Histone Acetyltransferase,HAT)结构域,因此PGC-1本身不具备这种内源性的乙酰化活性。有研究却发现PGC-1的N末端激活域能够招募具有HAT活性的其他蛋白,如类固醇受体辅激活因子 1(Steroid Receptor Coactivator 1,SRC-1)和CREB结合蛋白(CBP/p300),进而形成转录辅激活物复合体,同时研究发现SRC-1和CBP/p300与PGC-1的N末端的结合,需要依赖于PPARγ、NRF-1等转录因子结合于PGC-1第 200~400位之间的氨基酸序列。这与以前人们对辅激活因子的认识不同,PGC-1在与转录因子结合前处于一种相对的静止状态,当其与转录因子PPARγ、NRF-1结合后构象发生改变高度活化,继而招募SRC-1和CBP/p300至辅激活物复合体,最终导致其转录活性增加。PGC-1活性的开启环节在于同转录因子发生的分子对接 [26] 。
虽然现在关于转录辅调节因子的研究多集中于其参与调控转录起始的机制研究,但是大量的证据表明转录辅调节因子也能影响转录后的生物程序 [27] ,如mRNA剪接过程。PGC-1具有特征性的结构特征,能参与剪接加工、结合转录因子和辅激活因子。关于RNA特定的PGC-1结合靶点和mRNA加工和稳定作用的调控机制还有待进一步阐明。
磷酸化作用可能增加辅调节因子的酶活性,或者促进其他辅调节因子的趋聚,或者加速抑制性辅调节复合物的解聚。有研究发现p38活化的MAPK对PGC-1的 3个氨基酸残基(Thr262,Ser265,Thr298)产生了磷酸化作用。发生磷酸化作用后,PGC-1的稳定性和半衰期都有所延长。以前研究认为此区域在转录过程中起负调节作用,也在PGC-1与NRF-1和一些核激素受体发生分子对接时发挥重要作用。
综上所述,PGC-1影响特定靶基因活化的方式可以概括为以下 3个模型:(1)PGC-1能作为典型的转录辅激活因子结合于转录因子增强转录起始的效率。PGC-1也可能一定程度上增加转录因子的启动子特异性。(2) PGC-1影响特定基因表达,依靠参与RNA的加工过程,如延长、mRNA加帽、选择性剪接或增加mRNA的稳定性,而不是通过影响转录起始的环节。(3)综合上述两个模型,PGC-1动态参与特定基因表达的多重环节。
核呼吸因子(Nuclear Respiratory Factors,NRFs)是一类能调节线粒体呼吸链基因表达的转录因子,包括NRF-1与NRF-2。这两种因子在分子结构、作用机制等方面具有许多各自不同的特点。
NRF-1最早作为线粒体细胞色素C基因的一种转录因子被发现以后,又在许多编码线粒体呼吸链基因的启动子区发现了NRF-1的结合位点,证明它也参与了这些基因的转录调控。近年来的研究显示,NRF-1不仅参与线粒体基因的转录,还参与了线粒体基因的复制以及细胞发育过程中的某些相关基因的表达调节。
Gopalakrishnan等 [28] 首先详细研究了人类NRF-1基因的组成序列、转录和表达以及基因的染色体定位,发现人类NRF-1基因位于 7号染色体长臂 3区 1带内,含有 11个外显子,其中有两个外显子位于NRF-1基因 5′不翻译区(Untranslated Region,UTR),分别命名为 5 UTR-1和 5 UTR-2,其余 9个参与蛋白质编码,这些外显子的总长度约 65kb,该基因转录的启动子位于 5′UTR内。大鼠在正常生理状态下,体内各组织的NRF-1 mRNA维持在较低的水平,其中含量最高的是肺脏和睾丸组织,其次是心、肾和脑,而在肌肉和肝脏内含量最低。NRF-1作为一种核转录因子,在线粒体基因转录和复制的许多重要环节中发挥作用。近年来,已经在人类、大鼠、小鼠、牛等哺乳动物有关线粒体氧化磷酸化、线粒体DNA复制与转录、血红素合成、线粒体相关蛋白的翻译、输送、组装以及离子通道相关蛋白的基因上找到了NRF-1蛋白的识别位点。NRF-1中结合DNA的区域可以分成两个亚区,即含有核定位信号的结合DNA部分和含有磷酸化位点的丝氨酸残基部分。单个NRF-1分子的丝氨酸残基磷酸化后形成稳定的二聚体,与DNA上一段富含GC的回文序列结合后发挥作用 [29] 。
在有氧运动、肌肉收缩、甲状腺激素、内毒素、脂多糖等外界因素作用下,NRF-1mRNA表达量增加,蛋白分子结合DNA的能力增强,从而启动它所调节的靶基因转录,最终引起线粒体密度增加,线粒体呼吸链氧化磷酸化功能增强 [30] 。NRF-1在细胞生命活动中可能具有更为广泛的生物学功能,在一些与线粒体呼吸功能并无直接相关的基因,如在c-myc上发现了具有生物学活性的NRF-1位点,提示它可能参与凋亡的发生 [31] 。
在许多表达呼吸链所需成分的核基因中存在NRF-1识别位点。其一是氧化磷酸化作用所需组分,包括细胞色素C及 5个呼吸链复合体的部分亚基;其二是mtDNA转录和复制因子,如mtTFA和线粒体RNA加工内切核酸酶(Mitochondrial RNA Processing Endonuclease,MRP);三是亚铁红血素生物合成酶,如 5-ALAS(5-Aminolevulinate Synthase);四是与蛋白质输入和装配有关的组分,如Human Tom 20。NRF-1通过作用于这些基因对呼吸链的表达进行调节。
细胞色素C编码基因启动子的最大活性依赖于启动子中完好的NRF-1和cAMP反应结合蛋白(cAMP Response Element Binding Protein,CREB)结合位点。NRF-1和CREB通过磷酸化丝氨酸来激活细胞色素C编码基因启动子的活性。然而CREB的磷酸化作用是迅速和短暂的,而NRF-1的磷酸化作用发生较晚并更稳定 [32] 。
NRF-1的结合对人类mtTFA编码基因最关键启动子的激活起决定作用。mtTFA作为HMG基因家族的成员,是mtDNA转录和复制的必需因子,它被认为是脊椎动物线粒体基因表达最主要的激活调节因子。mtTFA在线粒体RNA聚合酶作用下,调节mtDNA轻、重两条链的转录与复制。人类mtTFA编码基因最关键启动子的激活高度依赖于NRF-1和NRF-2,而又以NRF-1的结合起决定作用 [33] 。因此,NRF-1既可直接调节核基因组编码的呼吸链亚基的表达,又可通过调节mtTFA而调节线粒体基因组的转录、翻译等,从而间接调节线粒体基因组编码的呼吸链亚基的表达。
NRF-2是一种具有称为ETS(E twenty-six)结构域的转录调节蛋白,由 α 、β、γ3个亚基组成。人类NRF-2分子含有一个 α 亚基、两个β亚基和两个γ亚基,其中 α 亚基含有ETS结构域即DNA结合区,由85个氨基酸组成,位于羧基末端,是一个经过修饰的螺旋-转角-螺旋结构,含有一个色氨酸三联体,能结合到靶基因的特定启动子区发挥调节线粒体呼吸链基因表达的作用。β亚基通过与 α 亚基形成二聚体后能加强NRF-2识别DNA并与之结合的能力,γ亚基可能与NRF-2分子转录调节的组织特异性有关 [34] 。
NRF-2也具有十分广泛的调控线粒体相关基因表达的作用,目前已经发现在线粒体呼吸链复合体Ⅱ,Ⅳ,V编码基因亚基以及mtTFA和血红素合成的限速酶以及与蛋白质合成、输送与组装、离子通道、穿梭机制相关基因序列上具有NRF-2的作用位点 [29] 。
人类线粒体转录因子A (Human Mitochondrial Transcription Factor A,h-mtTFA)是由核染色体 10q21编码的 25kD的蛋白,含两个HMG box结构域,属于典型的HMG box蛋白家族。通过对mtTFA的cDNA克隆分析,显示在其氨基末端含有一线粒体基质定位信号,之后为两个由约 80个氨基酸组成的HMG box DNA结合结构域,两个结构域之间被一短的连接区域分隔,其C末端尾区相对富含酸性氨基酸。突变和区域对换实验发现,HMG boxes对于非特异性的DNA结合特性是必需的,而C末端尾区对于特异性DNA结合和转录激活是绝对必需的,这表明mtTFA可能通过C末端尾区同其他与转录起始相关的因子发生相互作用。此外,mtTFA转录子在睾丸被进行 3种不同的剪切。在这 3种剪切体中,其中一种编码了缺乏线粒体靶向信号并定位于精母细胞和长形精子细胞核内的蛋白。因此,mtTFA基因可编码靶向线粒体和核的不同的蛋白质,这可能对于线粒体生物合成期间协调核和线粒体基因组的正确调控有着重要作用 [35] 。
目前已经鉴定得到了人、大鼠和小鼠mtTFA的启动子。序列分析显示它们启动子区含有 3个DNA结合基序:NRF-1、NRF-2和Spl。在培养的细胞中利用突变技术消除NRF-1和NRF-2在mtTFA启动子区的结合可以降低启动子的活性,表明NRF结合位点是mtTFA启动子活性的主要决定因素。
在哺乳动物中,mtTFA是线粒体DNA转录的关键激活子,其可结合至线粒体D-loop区内的重链和轻链启动子上的序列元件,并可在体外刺激以mtDNA为模板的转录。由于由轻链启动子起始的转录为重链的复制提供了必需的引物(线粒体DNA复制必需的第一步),故其同样在mtDNA复制中发挥作用。因此,mtTFA可能是决定线粒体拷贝数和转录活性的一个重要的控制点。mtTFA基因敲除小鼠表现为mtDNA缺失和氧化磷酸化不足,并在胚胎发育期死亡。由于真核细胞线粒体功能的维持需要两个生理上分离的基因组的基因产物的参与:一个包含在细胞器,另一个位于细胞核。所以,mtTFA的核起源为连接核和细胞器基因表达提供了一个非常有意义的研究对象 [36] 。
很多研究发现,mtTFA通过参与对线粒体转录的调控而对人寿命和疾病产生影响。在获得性甲状腺功能减退症中,mtTFA一直被认为是甲状腺激素作用的目标。在霍夫曼综合症(甲状腺功能减退性肌病)的病人中发现有mtTFA的减少和线粒体功能障碍。对老年人骨骼肌活检样本的研究发现,老年样本mtTFA的蛋白和mRNA水平分别比青年人样本增加了 2.6倍和 11倍,并发现mtTFA表达的增加同NRF-1对DNA结合亲和力增加和其mRNA水平升高有关。在对线粒体脑病病人的骨骼肌细胞的研究中也发现mtTFA处于异常水平,提示mtTFA可能同决定线粒体脑病功能性损伤的信号通路级联有关。
肌细胞增强因子-2(MEF-2)在肌纤维增生的基因表达调控中位于诸多信号传导通路下游,能激活很多重要的结构蛋白。MEF-2的结合位点在肌肉特异性基因调控区广泛存在,可以与多种调控因子结合形成复合体,多数与肌细胞发生相关的基因和蛋白质均与MEF-2关联。MEF-2结合到PGC-1 α 的启动子上使PGC-1激活 [37] 。MEF-2活性由CnA的去磷酸化作用激活 [38] 。组蛋白去乙酰酶基转移酶Ⅱ(HDACⅡ)是MEF-2的抑制剂,当其对钙信号产生反应的两个丝氨酸残基被磷酸化后,便从MEF-2上分离开来,使MEF-2游离 [39,40] 。CaMK能够磷酸化HDACⅡ,使磷酸化的HDACⅡ与MEF-2分离,游离的MEF-2在不同的通路中可被MAPK和PI3K等磷酸化激活其启动基因转录的活性。MEF-2将Cn和CaMK信号途径与PGC-1 α 的转录联接起来。PGC-1 α 上有 2个MEF-2的结合位点,可通过MEF-2的激活和HDAC的抑制调节PGC-1 α 的转录活性,诱导心肌线粒体的生物发生 [41] 。
转录因子YY1是诸多基因转录的负性调节因子,Spl是普遍存在的核因子,在维持“管家”基因的基本转录水平方面起关键作用,对特定的信号级联通路与应答这些信号的靶基因之间的调节也起重要介导作用 [42] 。Spl是既有正向又有反向调节作用的元件 [43] 。YY1与Spl相互作用,可形成复合体,与PGC-1、NRF-1等相互作用发挥其双向调节作用 [44] 。