
下载掌阅APP,畅读海量书库
立即打开



对藏品具有破坏力的昆虫(insect),其危害主要来自其取食、排遗及活动过程,对藏品造成的损坏包括形体损坏与污损。有些昆虫还有害藏品管理人员的健康,或给用户(参观或阅读者)带来滋扰。其实进入收藏单位建筑物内部的任何昆虫,都对藏品的安全具有威胁性,尽管某些昆虫对藏展物品并不构成直接危害,但如果它们死在建筑物内将引发继发性危害,因为它们会吸引其他昆虫或老鼠来取食,成为藏展环境虫害与鼠害的诱饵。
对于藏品管理人员而言,重要的是了解昆虫的特点,知晓昆虫的习性,识别昆虫的嗜好,以利于发现它们的存在,并将其拒之于藏展单位的建筑物之外,或通过适当的方法消灭侵入建筑物的昆虫。
了解昆虫的某些生物学特性与生态学特性,是保护藏品不为虫害的必备知识之一。
昆虫的生物学特性所包含的内容较多,以下仅对与危害藏品相关的某些生物学特性进行介绍,以便于对昆虫防治原理的理解。
1.1.1 形态特征
昆虫的身体可分为头、胸、腹三节。成虫通常有三对足和两对翅,头部有眼、一对触角和口器等,头部为昆虫的感觉和摄食中心。
(1)外骨骼与蜕皮
与脊椎动物不同的是,昆虫的身体并没有内骨骼的支持,昆虫身体外裹了一层由几丁质、蜡质层构成的轻巧且坚固的体壁,该体壁起着皮肤和骨骼的作用且会分节以利于运动。昆虫的几丁质外骨骼除保护内部器官外,还可减少虫体内水分蒸发以适应陆地上的干燥环境。昆虫坚硬的体壁使其生长受到限制,蜕皮(ecdysis)成为昆虫发育的一种生理现象。昆虫蜕皮时,体形可能骤变。正常情况下,每蜕一次皮,昆虫不仅躯体增大,其内部各器官也得到进一步的发育。
昆虫蜕皮的次数因种类而异,大部分较高等的昆虫一生蜕皮的次数比较固定,常见为3~6次,低等的昆虫如衣鱼,一生不断蜕皮,次数可达60次之多。
 环境因素(如环境温度与食物)的改变有可能改变昆虫蜕皮的次数。通常环境温度升高可能增加蜕皮次数,但也有一些昆虫在温度升高时蜕皮次数反而减少。营养不足也有可能增加昆虫的蜕皮次数,如皮蠹科昆虫(Dermestidae)与幕衣蛾(
Tineola bisselliella
)等的幼虫,在饥饿或太干燥时,会大大增加蜕皮次数,例如幕衣蛾的蜕皮次数可由4次增加到40次之多,发育期则延长。不过,营养不良带来的蜕皮次数增加,不仅不会使虫体增大,反而使之越变越小。
环境因素(如环境温度与食物)的改变有可能改变昆虫蜕皮的次数。通常环境温度升高可能增加蜕皮次数,但也有一些昆虫在温度升高时蜕皮次数反而减少。营养不足也有可能增加昆虫的蜕皮次数,如皮蠹科昆虫(Dermestidae)与幕衣蛾(
Tineola bisselliella
)等的幼虫,在饥饿或太干燥时,会大大增加蜕皮次数,例如幕衣蛾的蜕皮次数可由4次增加到40次之多,发育期则延长。不过,营养不良带来的蜕皮次数增加,不仅不会使虫体增大,反而使之越变越小。
由于蜕皮是昆虫生命中不可少的环节,依此规律有助于发现虫害,如在虫害发生处或在其周围有可能发现虫蜕(即昆虫脱下的皮),但也有些昆虫会吃掉自身的虫蜕。
接近蜕皮时,幼虫会逐步停止取食并排出排遗物,吐少量的丝以固定虫体,静止不动地进入蜕皮过程。故在书页卷曲处、粘结处常常可能发现休眠状的幼虫。
(2)形体小
昆虫体型较小,成虫的长度都是以毫米(mm)进行计算,例如:白腹皮蠹成虫6~10 mm,火腿皮蠹成虫7~10 mm,褐粉蠹成虫5~6 mm,黄蛛甲成虫3~4.5 mm,报死窃蠹成虫4~7 mm,等等。
躯体小给昆虫的生存带来很多优势,例如所需的生活空间就小,更利于隐藏、躲避侵害。同时,也便于与寄主一起移动而广为传播扩散,由此使其流动性强。如危害藏品的昆虫可寄居在藏品、装具中,随藏品巡展、交流等人类活动而全球蔓延。
体小的昆虫对环境的控制能力小,为了生存,其对环境的适应能力就强。昆虫最适宜在温暖的环境中生存,全球气候变暖,可能会对昆虫种群产生许多影响,如使昆虫跨地域生存能力加强,这些都使得很多危害藏品的昆虫生存的地理区域性界线不明显。有证据表明,危害藏品的昆虫正在向新的地域扩散,这需要我们在虫害分布与风险评估上考虑全球气候变化。也正因为这个,本书附录8危害藏品的昆虫分布列出了在世界其他地区发现的危害藏品的昆虫,以便我国藏品管理人员知晓与警惕。
1.1.2 昆虫的某些器官
昆虫的器官较多,这里仅就与昆虫危害与消杀相关的器官进行介绍。
(1)感觉器官
昆虫体型虽小但感官却很发达,可以看到人眼看不到的光线,听到人耳听不到的声音,嗅到不同的气味。
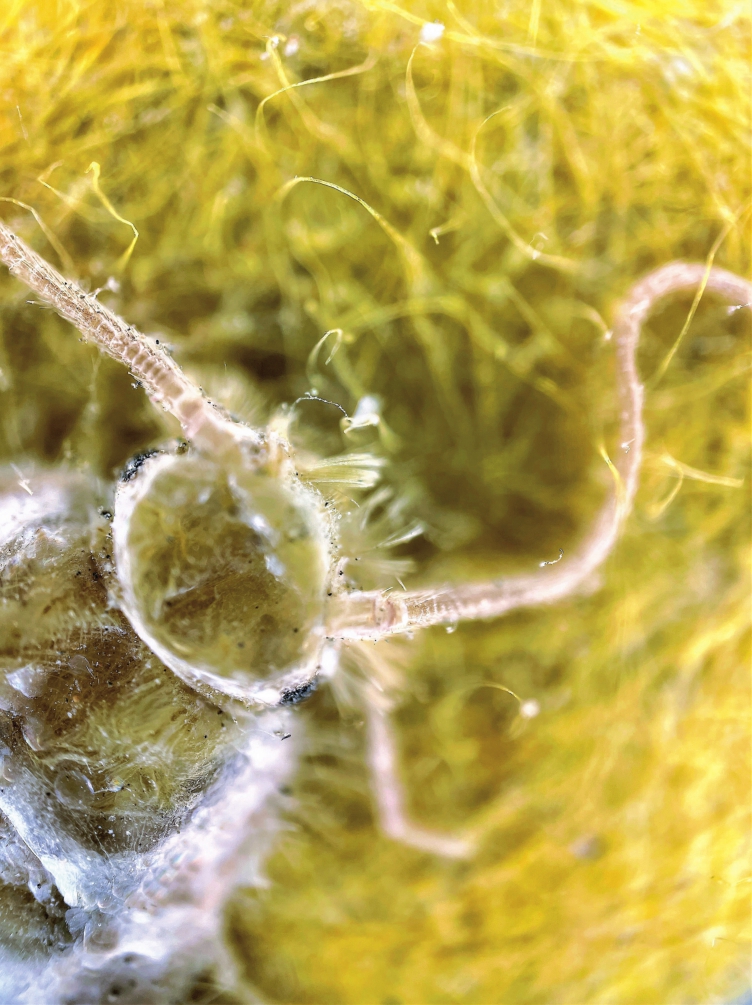
图1-1 昆虫的头部
摄影:广州文保文化传播有限公司
昆虫的感觉器官主要集中在头部,头部的多个感觉器官可获取与处理外界信息。例如,其嗅觉就分布在触角与口须。长在昆虫头部前上方的触角随昆虫种类不同而形状各异,有的细长像一对鞭子,有的则像两把刷子,有的更像两把短锤,等等。不同形状的触角上分布着许多感觉器和嗅觉器,除了能够感知物体与气流,更能嗅到各种气味,甚至百米之外的同类的气味。在昆虫口部下方的两对短小的口须与触角一样分布了许多能感知气味的嗅觉器,使它们能够辨别气味。昆虫就是依据这些嗅觉器官,辨认不同藏品发出的不同气味信息,准确地找到最喜爱吃的藏品。由此,这类藏品也就成为最可能受害的高危藏品。
除头部外,昆虫的感觉器官可能位于昆虫体内的几乎任何位置。例如,其外骨骼也具有许多感知器官,可检测光线、压力、声音、温度、气流和气味。又如,其触角除可感知外,有时还会传递气味信息等。
(2)呼吸器官
氧气是维持动物生命的最重要因子之一,昆虫是需要吸入氧气呼出二氧化碳的。与人不同的是昆虫没有肺,它是通过胸部和腹部两侧的圆形小孔(即气门)进行呼吸的,气门连着气管,气管又分支成许多微气管,通向昆虫身体的各个部位。昆虫依靠腹部的一张一缩,通过气门、气管进行呼吸,使体内维持一定的氧气浓度。正常情况下,昆虫的呼吸数基本是恒定的,这是由气门开闭的时间来控制的。体壁上气门的开合控制,也有效地减少了因蒸腾作用而造成的虫体水分流失。
遇到外界环境改变,如空气中氧气浓度降低,为了维持体内氧气浓度,昆虫会加速呼吸,此时昆虫会延长气门打开的时间,直到可吸入所需的氧气。昆虫呼吸越急促,气门打开的次数越频繁,虫体水分散发就会越多。若气门被迫持续打开,而外界无法补充水分给虫体,昆虫就会干萎而死。低氧气调灭虫后的死虫多为干瘪状,就是其失水而死所致。
(3)取食器官
昆虫的取食器官称为口器(trophi),位于昆虫头部。不同的昆虫因为取食方式不同,其口器类型也不同。
在很多情况下,对藏品造成最大破坏的是昆虫的幼虫,但也有例外,例如衣鱼,其若虫与成虫都具有取食藏品的能力。这些具有取食藏品能力的昆虫,其口器都是咀嚼式口器。
咀嚼式口器是最原始的口器类型,其主要特点是具有发达而坚硬的上颚,用来咬碎与嚼碎固体食物,给物品带来直接伤害。如钻蛀性昆虫常将器物蛀成隧道或孔洞;咬碎性昆虫的幼虫能将被害物咬成片状,直到被害物完全被毁坏;侵蚀性昆虫啃食物品表面,留下虫洞与抓痕(更多相关内容见:2.5依取食特点分类)。危害物品的昆虫利用其发达的咀嚼式口器咬啮或蛀穿藏品的同时,留下各种形状的咬食痕迹(食痕),这为辨别昆虫的种类提供了一定的证据。
昆虫口器的构造还会随昆虫的发育有所变化,如危害藏品的鳞翅目昆虫的幼虫,其口器多数为咀嚼式口器,待其发育到成虫则会转变为虹吸式口器。又如危害仓储物品的鞘翅目昆虫的成虫与幼虫都是咀嚼式口器,但其成虫口器退化,不能再取食固体食物,基本丧失了对物品的危害能力,但钻蛀性害虫为离开物品而咬的出口孔仍然会对物品造成伤害。
1.1.3 繁殖能力
昆虫是地球上数量最多的动物群体,直到21世纪初,人类已知的昆虫已经超过100万种
,但仍有许多种类尚待被发现。环境的恶劣和种群繁衍的需求促使昆虫具有强大的繁殖能力,有些昆虫具有一次受孕终身产卵的能力,如鞘翅目的所有甲虫以及蟑螂、蟋蟀等。
大多数危害藏品的昆虫都具有繁殖力强的特点,如在25℃的标本馆内,烟草甲每天可产10~20粒卵,黑毛皮蠹每头雌虫可产450~900粒卵。
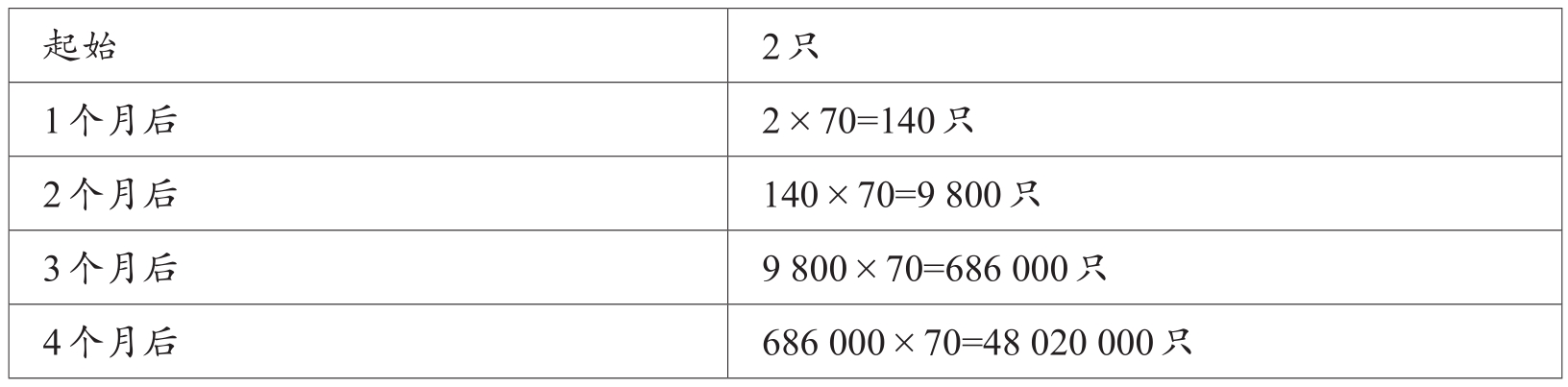
有学者指出,谷蠹的增殖因子是70,即在最佳条件下,其后代每月数量都在呈70倍的增加。
表1-1 谷蠹后代数量的繁衍
昆虫除有性生殖外,亦有孤雌生殖、幼体生殖等现象,即卵不经过受精就能发育成新的个体。
以上特点,使得危害藏品的昆虫一旦进入收藏单位,即可从少数个体发展到泛滥成灾的程度,管理人员一旦发现虫情,就必须尽快采取治理措施。
1.1.4 生活习性
昆虫的习性包括昆虫的活动及行为,了解不同种昆虫的不同习性有利于虫害的防治。昆虫种类繁多、形态各异,遍布全球,所以生活习性是大不相同的,以下介绍的仅是危害藏品的昆虫的主要共性,相关昆虫的生活个性见:4.重要害虫。
1.1.4.1 食性
昆虫的生长与发育离不开食物,在食物营养丰富与充足的情况下,虫口密度才能得到快速增长。但不同种类的昆虫对自身的食料有明显的选择性和适应性。食性(feeding habit)是指昆虫对食物的一定要求,即昆虫取食习性、取食范围及取食偏好。昆虫对食物的选择性与昆虫的口器、消化系统和共生体相关。
不同种类的昆虫具有不同的口器与消化系统,因此对食物具有一定的要求与选择。例如尽管干燥的植物种子和被淀粉糊处理过的纸都含有大量淀粉,但象鼻虫更容易攻击干种子,而衣鱼则优先攻击有淀粉糊的纸。出现这一区别的主要原因是二者的口器结构差别太大。尽管象鼻虫与衣鱼都可以消化淀粉,但二者口器结构不同。象鼻虫头前有个如象鼻一般的长长口器,可以刺穿植物组织进行取食并在里面产卵,但衣鱼的口器很原始,虽然露在头外,但只能在材质表面啃咬,难以取食干种子,故优先攻击纸张上的淀粉了。
不同种类的昆虫可能具有不同的消化系统与共生体,使得它们对不同食物具有的消化能力不同。共生体是指不同物种的生物之间的紧密且长期的相互作用。例如,有些昆虫肠道内的微生物可将纤维素转化为可用的糖类并消化,有些昆虫肠道内就没有这类共生体,有这类共生体的昆虫就具有取食植物性纤维的能力。
昆虫的以上特点决定了它们的危害对象大多具有专化性,即攻击对象是具有一定特点的材质。了解主要昆虫的食性,便于有针对性地采取防控措施。
依据昆虫取食的偏好,危害藏品的昆虫可分为4大类,即危害植物源性纤维的食植昆虫(herbivore)、危害动物源性纤维的食肉昆虫(carnivore)、杂食性昆虫(omnivore)与腐食性昆虫(saprophagous)。其中杂食性昆虫为不挑食的昆虫,其他类型的昆虫都有其最优先取食的对象(见2.4依昆虫食性分类)。利用昆虫的该特点,可识别昆虫以及防治昆虫。
需要说明的是,尽管昆虫取食具有专化喜好性,但并非绝对不取食其他类型的食物,只是在食物具有多种选择时,才会执行优先取食的选择。例如,喜食动物源性纤维的昆虫,只在同时存在植物源性纤维、动物源性纤维时,才会优先攻击动物源性纤维,但并非严格拒食植物源性纤维。又如一些喜爱以干燥植物材料为食的昆虫,如蛛甲、烟草甲等,其幼虫不仅会危害植物标本,也能够侵扰基于角蛋白或几丁质的材料,只是当一个区域内有多种材质的物品时,它们更喜欢以植物源性纤维为食。再如药材甲的食性很杂,动植物干品均可取食,但它更爱取食植物源性干品,故为中草药与植物标本的重要害虫。
昆虫取食的专化性除直接威胁到不同材质藏品对昆虫的敏感性外,也直接影响到昆虫自身的生长发育。若能取食到最喜欢吃的食物,昆虫就发育快,死亡率低,生殖力强。有些昆虫尽管属于不挑食的杂食性昆虫,但不同的食物仍然可以影响杂食性昆虫的发育速度、存活率、生殖率及滞育等各方面。
有研究表明,昆虫消耗食物的速度还受到环境条件、食物本身性质(如食物的时代、营养价值和特殊成分)的强烈影响。
1.1.4.2 释放气味信息
昆虫个体可向体外释放非常特殊的挥发性化学物质——信息素,同物种的其他个体通过嗅觉器官可以感知这类特殊的气味信息,它是同物种不同昆虫间沟通信息的通讯工具。
昆虫至少可以释放5种信息素:报警信息素(alarm pheromones)是向同种昆虫发出警告或呼叫求助并发动攻击的信息素;踪迹信息素(trail pheromones)是被社会性昆虫(如白蚁)用来记录自己踪迹,引导该物种其他成员跟随的信息;性信息素(sex pheromones)是昆虫散布求偶的信息,因为大多数成虫的寿命都很短,所以使用性信息素来迅速吸引配偶并排斥其他物种;聚集信息素(aggregation pheromones)是告知同一物种的个体到此群聚的信息,个体聚积越多,该位置的信息素的浓度就越高,如蟑螂就是利用该信息素聚集活动的;当昆虫的个体密度达到一定程度时,会释放分散信息素(dispersing pheromones)以保持个体之间的距离。
人类利用昆虫这类行为沟通工具,研制出某些信息素(如性信息素)用于诱捕昆虫,以检测目标昆虫的存在,确定其丰度或进行控制。
1.1.4.3 抗逆性强
抗逆性是昆虫在逆境中得以生存的关键特征。
当昆虫遇到不良环境时(如高温或低温,食物短缺),它可以通过休眠(dormancy)来适应外界环境以求得长期存活。
在不宜昆虫生长发育的恶劣环境中,昆虫会通过暂时停止生长发育的方式延缓生命,这种现象即为休眠。一旦环境适宜,昆虫会从休眠状态苏醒,恢复其生长发育。休眠的昆虫呼吸和新陈代谢显著降低,并且长时间不食也不动,以此承受正常生物无法生存的严酷低温、低氧、无补给的恶劣环境。因此,无论是冷冻灭虫还是低氧灭虫,都需要在灭虫结束后在隔离室观察一段时期,避免其在入库的正常环境中苏醒,再次危害藏品。
例如,花斑皮蠹的抗逆性很强,在温度为38~40℃,储藏物含水量8%~10%,RH≤50%的条件下,均能正常发育和繁殖。
百怪皮蠹对高温、低温的抵抗力很强,成虫在0℃和0℃以下可存活37天,幼虫在-6.7~3.9℃下可存活67天,-15~12.2℃下可存活12天。
在缺乏食物情况下,花斑皮蠹休眠幼虫可耐饥8年不死。
又如,日本蛛甲在-5℃下能存活164天;东方蜚蠊在0℃下可生活91天;黑毛皮蠹成虫在40℃左右仍能继续繁殖,其幼虫在-3.9℃时能存活198天;异斑皮蠹耐饥能力强,可以5年不食。
在环境不利时,有些昆虫可以通过蜕皮缩小虫体以延长其生命,如家扁天牛(
Euryphagus antennata
)、衣蛾(
Tinea
)、谷斑皮蠹(
Trogoderma granarium
)。
在蜕皮过程中,蜕皮液可将虫皮内营养物质消化吸收,以此作为虫体继续生存的物质能量来源。
昆虫的抗性不仅体现在在不宜生存的藏展环境中的强大生命力,也体现在灭虫过程中的不易杀灭性,这种抗逆性与昆虫的物种以及其生命阶段相关。
1.1.4.4 变态发育
生命周期(life cycle)是指昆虫经历孵化、发育、死亡的全过程,通常这一过程也称为一个世代(generation)。昆虫在其生命周期不同发育阶段表现出不同的形态、结构、功能和生活习性的变化被称为变态(metamorphosis)。
昆虫因种类不同,生活情形各异,变态也不一致。昆虫的变态可以划分为两大类:完全变态(complete metamorphosis)与不完全变态(incomplete metamorphosis)。
(1)完全变态
完全变态的昆虫在其发育过程中要经历4个阶段(虫期)的变化,即卵、幼虫、蛹、成虫。在其生活史中,所有的生长都发生在幼虫期,其卵、蛹和成虫都不会长大。完全变态的昆虫的成虫在适当的地点、适合的材料上产卵,微小的幼虫(larvae)从卵中孵化出来,它可能立即钻入藏品材质中,在不断蛀蚀藏品中长大;有的微小幼虫并不立即钻入藏品中,而是在藏品表面到处觅食而侵扰藏品材质。随着幼虫不断长大并老熟后,就会经过一个吐丝结茧并在茧内化蛹的过程,在化蛹期间幼虫组织完全改变而转化为成虫构造。为了降低蛹被其他昆虫捕食的几率,化蛹可能发生在远离其食物处或其他较为隐蔽的角落。成虫一旦羽化,性器官就成熟,不需要取食即可交尾、产卵,这时成虫口器一般都会退化,寿命很短。有的成虫口器发生变化不再具有取食固体物质的能力,如鳞翅目的蛾类,其口器变为虹吸式,只有吸食花蜜的能力。
由上可见,完全变态昆虫的幼虫是直接危害物品的主要虫期,成虫与幼虫具有不同的食性,并生存在不同空间。大多数成虫不再具有对物品的直接危害能力,而成为昆虫的繁殖工具。对仓储物品危害最大的鞘翅目、鳞翅目均属于完全变态。需要注意的是,有些成虫尽管不再取食物品,但若其在物品内化蛹,则会咬破物品而离开,这时物品也会受到危害,例如留下不同形态的羽化孔(emergence hole)。羽化孔一般是指钻蛀性昆虫由蛹变为成虫,从被害物品内部的蛹室爬出时在物品表面留下的孔洞,在国内又称为成虫出口孔(exit hole)、成虫出现孔等,本书统一称为“出口孔”,以更利于非专业人员理解。
(2)不完全变态
不完全变态昆虫与完全变态昆虫不同的是,其发育过程中缺失化蛹阶段,其生长发育只经过3个阶段,即卵、若虫期、成虫期。昆虫的卵经孵化而成的“幼虫”被称为若虫(nymph),体态与成虫相似,但异于成虫。不完全变态的过程比较缓慢,若虫在生长过程中要经历多次蜕变,才能完全发育成成虫。
从卵孵化出的若虫与成虫形态基本相同,只是在生长发育过程中个体增大、性器官逐渐成熟,有的在成虫期还会继续蜕皮,如衣鱼;有些若虫与成虫相比,不仅性器官尚未成熟,其翅也尚未发育完全,如蟋蟀、书虱。与完全变态昆虫不同的是,不完全变态昆虫的若虫与成虫的习性完全相同,如不仅食性相同,其栖息地或寄主也相同。如衣鱼,其若虫和成虫生活在同一空间,都以纸张、淀粉与纺织品为食。由此,不完全变态昆虫的幼虫与成虫都具有直接危害藏品的能力。
1.1.4.5 假死性
不少昆虫具有假死性(play dead),假死性的昆虫不同于休眠的昆虫。昆虫的假死性实际上是一种很简单的刺激反应,当遇到外界惊扰,其神经就会发出信号,使昆虫的浑身肌肉收缩起来,暂时停止活动或自动坠落,好像死去一样。过片刻后,它又爬行或起飞。烟草甲、档案窃蠹、竹蠹、拟裸蛛甲等危害藏品的昆虫,不少具有假死性。
昆虫的生命活动受很多环境因素的影响,如光、空气流动、食物、温度与相对湿度,了解昆虫与周围的环境的关系,是防治昆虫的基础。
光照变化可以极大地影响昆虫的活动和繁殖力,如飞蛾在黎明和黄昏时最活跃,人造光可以大大限制飞蛾的飞行活动和繁殖力
。空气流通可以降低气温和相对湿度,从而对昆虫的体温和水分产生影响,昆虫最喜欢生活在空气不流通处。
食物直接影响昆虫的生长、发育、繁殖和寿命。若食物数量足、质量高,昆虫生长发育就快,自然死亡率低,生殖率高;反之,则生长慢,其发育和生殖均受到影响,饥饿同样也会引起昆虫个体大量死亡。某些昆虫成虫期有取食补充营养的特点(如不完全变态的成虫),若得不到营养补充,则产卵甚少或不产卵,寿命也会缩短。
除以上因素外,与昆虫的生命活动关系非常密切的生态因子主要是环境温度与相对湿度,特别是温度的影响最大。每种昆虫都有适于其发育的最佳温度和最佳相对湿度,在不利其发育的条件下,其发育期会延长。昆虫的这些特性,对于有效控制昆虫至关重要。
1.2.1 温度
昆虫属变温动物,其体温的高低随环境温度变化而变化,温度是决定昆虫生长发育速率的最重要的因子。
昆虫的新陈代谢是在各种酶和激素的作用下进行的一系列生化反应,其生化反应的速度与虫体温度有关,其体温的变化可直接加速或抑制虫体的代谢过程。由于虫体的热量来源于环境,故昆虫的生长、发育的速度明显受制于环境温度。有研究指出,低于15℃的环境温度对减缓或阻止危害藏品的昆虫生长、摄食和繁殖非常有效,有时可以作为抑制虫害的临时措施。
图1-2是药材甲幼虫发育的速率与温度的关系图
[1]
,图1-3是昆虫发育率与温度的关系图
。由图1-2可见,昆虫幼虫的发育速率随环境温度升高而加快。图1-3类似于抛物线的曲线说明,在较低的环境温度下,昆虫基本没有发育迹象;随环境温度上升,昆虫发育速率增加并达到最大值,若温度继续增加,其发育速率迅速下降并趋近于零。
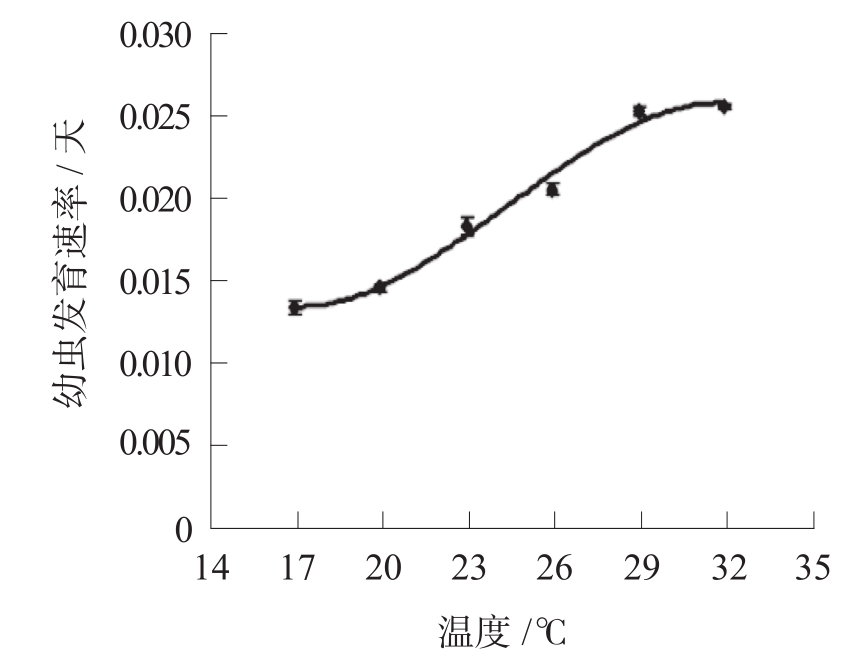
图1-2 药材甲幼虫发育速率与温度的关系
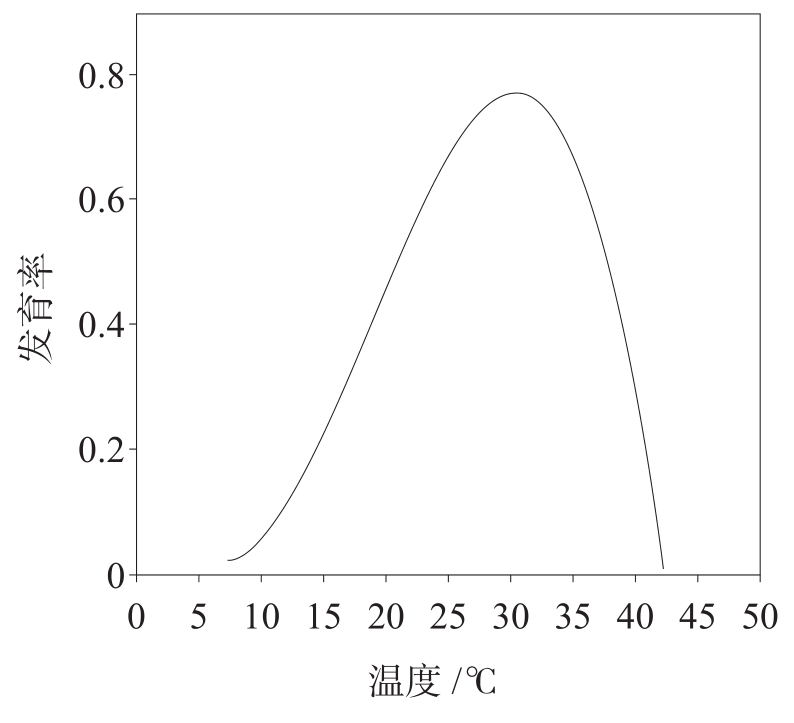
图1-3 昆虫发育率与温度的关系
尽管环境温度的升高提高了昆虫体温并促进了虫体代谢,但其温度上升的峰值不是无限的。过高的虫体温度可以致死昆虫,如虫体温度过高会导致昆虫体内水分剧烈汽化并蒸发而脱水致死,或是高温导致虫体蛋白质凝固变性,或高温破坏了虫体的细胞线粒体,甚至高温也可以抑制虫体内酶与激素的活性,加速虫体各生理过程的不协调,轻者会抑制昆虫的发育或给昆虫留下一些后遗作用,如引起成虫发育不全,体小,翅膀不能正常展开,或是不孕卵数量增多等;重者则造成昆虫死亡率增加或直接导致昆虫死亡。
较低的环境温度不仅会抑制昆虫的生长与繁殖,同样也会加速昆虫的死亡。在不太低的低温下(如0℃左右或稍高),一些耐寒性弱的昆虫体内养分会逐渐缺失,出现生理失调,尽管昆虫可采用休眠以抵抗,但若在较短时间内不能恢复到正常温度就可能死亡。
有研究将仓储昆虫可正常进行生命活动的温度范围称为适温区或有效温区,并参照大多数昆虫发育的速度,将适温区再划分为三个温区,即高适温区(33~35℃)、最适温区(25~33℃)与低适温区(13~25℃)。最适温区是大多数仓储昆虫生长和繁殖的最佳温度,大多数仓储昆虫在高适温区或低适温区是能够完成发育并产生后代的,但低于13℃或高于35℃,仓储昆虫会逐步走向死亡。温度越极端,它们死得越快。在-20℃或55℃的温度下,大多数仓储昆虫几分钟内就会死亡。致死温度的差别很大,这取决于物种、发育阶段、昆虫对环境的适应和环境的相对湿度等。
表1-2是仓储昆虫对温度反应概率统计的研究
,它更详细地展示了仓储昆虫在不同环境温度内的反应。
表1-2 仓储昆虫对温度反应的概率统计
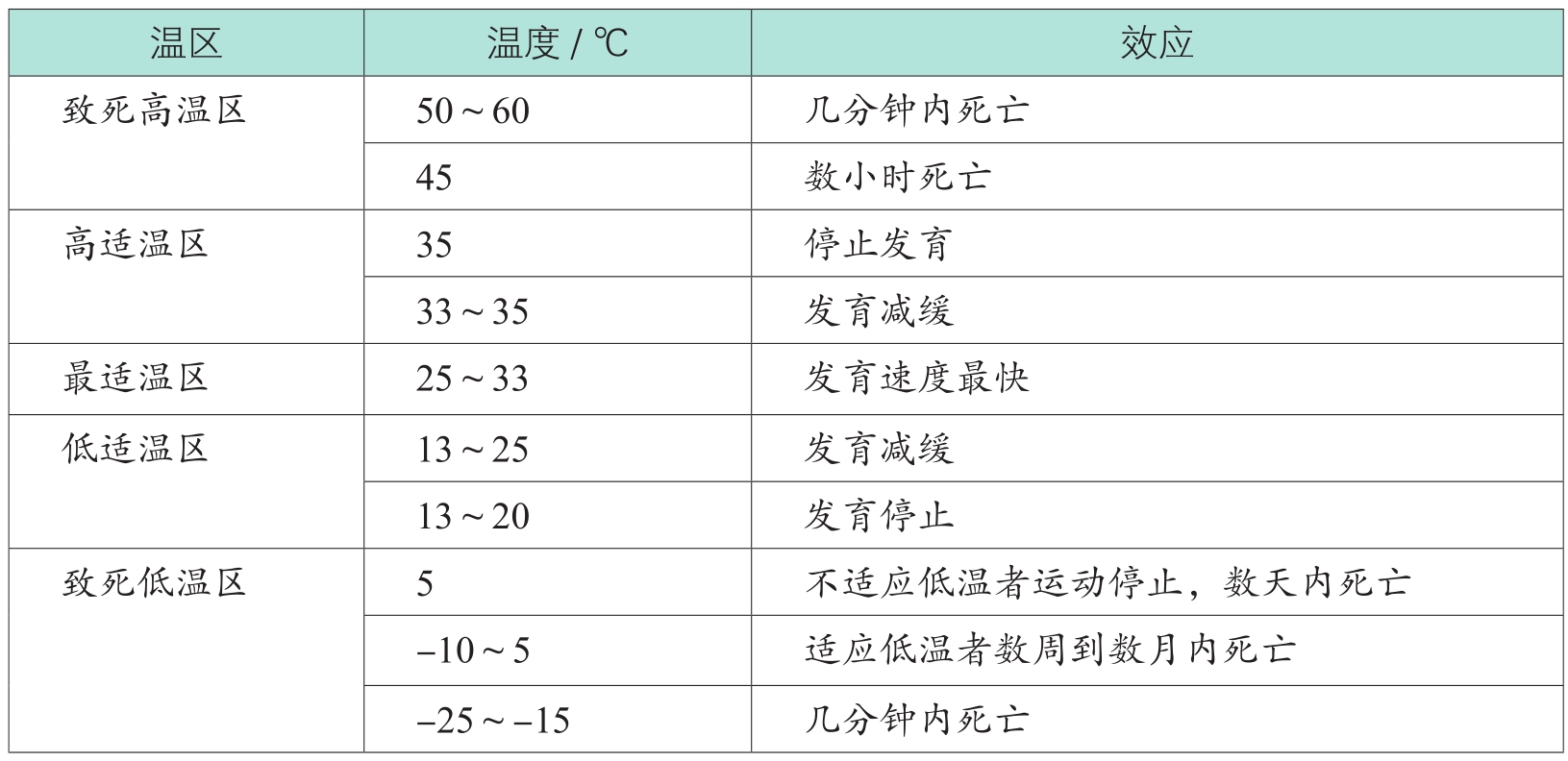
每种昆虫或同种昆虫的不同虫期(如卵、幼虫、蛹、成虫)都有自身发育的最低温度和最高温度的临界值(或称为阈值温度),超出最高温度或低于最低温度,昆虫发育就会停滞,其死亡率增大。但在达到致死温度之前,昆虫不会死亡。
有研究指出,危害藏品的常见昆虫平均发育阈值(LDT)顺序为:螨(6.8℃)和双翅目(8.1℃)的发育阈值较低,鳞翅目(11.3℃)和啮虫目(13.8℃)次之,鞘翅目(14℃)和蜚蠊目(15℃)的发育阈值较高。
昆虫在致死高温区与致死低温区的死亡,与恶劣温度的程度及其持续的时间均有关系。在致死高温区,昆虫会出现热昏迷状态,并随时间的延续而死亡。在高温下一时尚未杀死的昆虫,高温对该虫的生命与繁殖均会产生不利影响,如存活率显著降低,卵孵化率降低。
在致死低温区,大多数昆虫体内原生质析出水分结冰,不断扩大的冰晶可使原生质遭机械损伤、脱水,生理结构受到破坏,细胞膜破损,从而引起组织或细胞内部产生不可复原的变化而引起昆虫死亡。尽管有些昆虫是可以耐受结冰的,即组织中有冰形成时也能存活,但这种耐受性是相对的,随着温度的持续下降和处于冻结状态的时间延长,其存活率同样下降,所有的仓储昆虫在冷冻时都会死亡。
可见,利用低温杀灭昆虫既要考虑到低温的强度也要强调低温持续的时间。一般说来,在致死低温区,温度越低,昆虫死亡越快,杀灭昆虫所需的时间越短。但处于冷昏迷的昆虫一旦回到了正常温度,其生命活力就会恢复,因此灭虫低温的持续时间十分关键。
从昆虫的抗逆性强弱分析,昆虫耐寒性较耐热性强,即昆虫忍受低温强度和持续时间的能力相对较强。这与昆虫体内含有大量糖、脂肪、蛋白质等物质,与原生质形成一定的有机结构,使其体液可以忍受0℃以下的一定低温而不结冰有关。
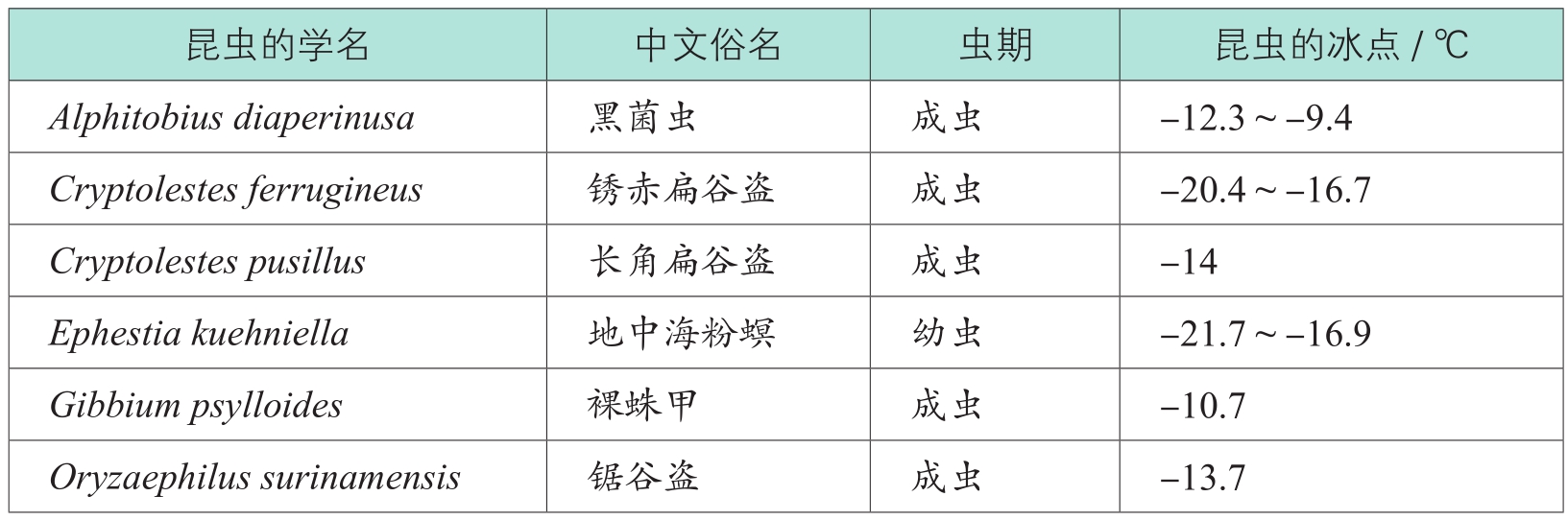
昆虫耐寒性的划分主要是依据其过冷却点的高低,过冷却点(supercooling points,SCP)是昆虫体液开始结冰时的体温,它代表着该类昆虫能够存活的温度下限。昆虫的体液降到冰点以下不一定都会结冰,只有在环境温度达到其过冷却点时,其体液才开始结冰,这个现象对于冷冻灭虫相当重要。表1-3列出了某些昆虫体液结冰的温度。
表1-3 某些昆虫的冰点
续表
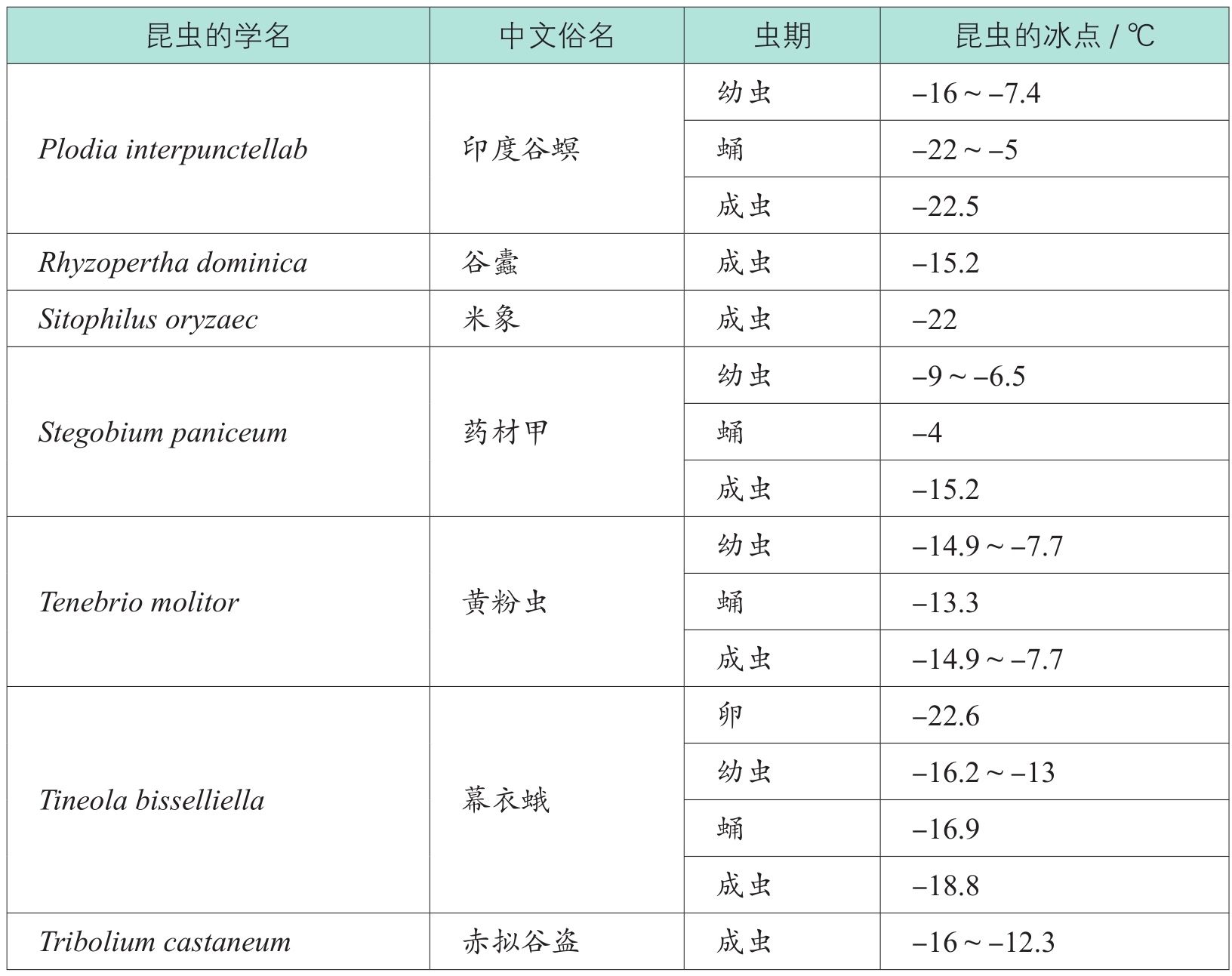
由表1-3可见,不仅不同昆虫的过冷却点不同,即使同一昆虫的不同发育期的过冷却点也不一样。一般说来,昆虫的过冷却点为-22~-4℃
,随昆虫的物种、体重、发育阶段、营养状况的差异、暴露于低温时间的长短以及遗传因素、气候等的变化而变化
。通常情况下,越冬虫态的耐寒性最强,休眠或滞育虫期对低温的耐受能力较强,而发育中的虫态耐寒能力较差。例如,蛾类基本上是以蛹和老熟幼虫越冬的,这两个虫态脂肪含量高,较耐寒,尤以蛹耐寒性最强。故甜菜夜蛾不同虫态耐寒性由弱至强的顺序是:卵、成虫、幼虫和蛹。
又如,西藏飞蝗四川种群以卵越冬,卵期的过冷却点显著低于其他虫态。
即使同一虫种在同一个虫期内,耐寒性也不同。如在飞蝗的卵中,因胚胎的发育阶段不同,耐寒性不同,初产的卵耐寒性显著高于胚转期和孵化前的卵。
家蚕完好卵要比受伤卵的过冷却点低,滞育卵和非滞育卵的过冷却点也有显著差异。
昆虫的耐寒性受环境影响很大,如缺氧条件可以提高昆虫的耐寒性,故高纬度、高海拔地区的昆虫种类或种群通常具有更强的耐寒能力。又如,家蝇蛹在无氧条件下处理10分钟,就能使将其在-7℃处理2小时的存活率从0提高到25.3%。
昆虫在被冷冻前若曾经经历过一段时期的低温,就可提高该昆虫的耐寒性,这也就是越冬的昆虫的抗寒性强的原因。
1.2.2 相对湿度
相对湿度(relative humidity, RH)。人类能够忍受广泛的相对湿度,但在45%~65%的范围内最舒服。一般来说,温度较低或较干燥的环境对昆虫的吸引力小。
大多数昆虫喜欢选择在湿度较高的环境内生活,如家具窃蠹、档案窃蠹,有人发现凡是捕获到档案窃蠹的库房,库房环境湿度都超过了55%。
有些昆虫对潮湿环境有强烈的偏爱与依赖,只能在潮湿环境下生存,例如衣鱼与书虱等,这类昆虫只有在湿度超70%以上的环境才可能快速繁殖并引起严重的问题(更多见:2.3.1喜湿昆虫或卫生昆虫)。
也有不少昆虫对环境湿度有较大的容忍度,在非常干燥或潮湿的环境中都可以存活很长时间,例如药材甲发育的最佳湿度为60%~90%。
尽管这类昆虫可以在接近60%的相对湿度下生存,但在潮湿的环境下更能茁壮成长。
[2]
尽管就环境湿度而言,每种昆虫的需求不同,但整体而言,大多数昆虫的生长、发育的适宜湿度为70%~90%
,因为相对湿度会影响到昆虫的生长、发育和繁殖能力。例如档案窃蠹,50%以下的相对湿度不利于卵的孵化,在36%的相对湿度条件下,卵不孵化。
家具窃蠹或某些蠹食木制品的昆虫,只有环境相对湿度在55%以上才能成功完成其生命周期。
相对湿度过高或过低都可以抑制昆虫的新陈代谢而使其发育延迟,昆虫卵的孵化、蜕皮、化蛹、羽化,一般都要求较高的相对湿度。在大多数情况下,控制环境的相对湿度对虫害具有一定的抑制作用。但这只是一般规律,不同种类的昆虫、同种昆虫的不同发育阶段都有可能对相对湿度具有不同的需求(更多见:4.重要害虫)。
一般情况下,降低藏展场所的相对湿度,可以抑制昆虫的发生与发育。这就是为什么在选择藏展环境的相对湿度时,在考虑藏品材质的前提下,应尽量选择材质可以忍受的相对湿度下限作为藏展环境的相对湿度,如保存纸质藏品的相对湿度为50%~60%,选择50%作为其保存环境的相对湿度更不利于昆虫的滋生。
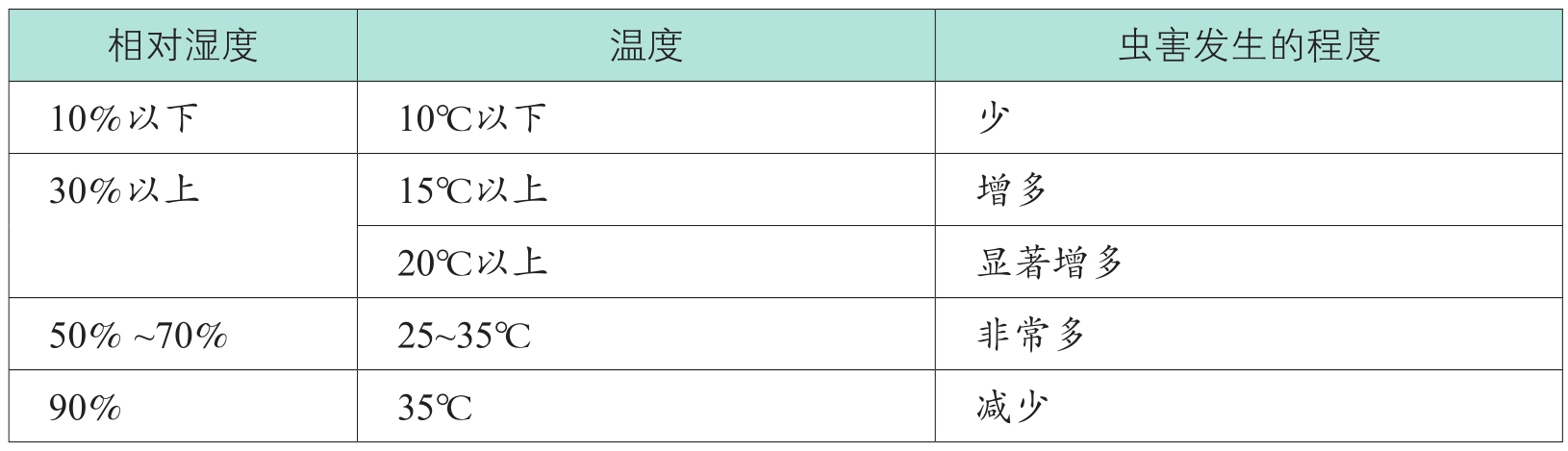
高温与高湿是任何昆虫都喜欢的生存环境,表1-4
呈现了这一事实。
表1-4 环境温湿度与虫害发生的程度
[1] 李灿,金道超,柳琼友,等.温度对药材甲( Stegobium Paniceum L. )实验种群发育和繁殖参数的影响[J].生态学报,2007(8):3532-3535.
[2] Opit G P,Throne J E. Effect of Diet on Population Growth of Psocids Lepinotus reticulates and Liposcelis entomophila [J]. Econ Entomol, 2008(2): 616-622.