
下载掌阅APP,畅读海量书库
立即打开



本书的主题是我们将称之为“基因—文化协同进化”的东西。乍一看这一措辞可能好像暗示了一种未必会发生又或许不可能的过程的耦合。但这并不是实情。生物学进化与文化进化之间的联合是一种逻辑可能性,其探索已成为一个越来越明显的重大的智力挑战。许多哲学家与科学家仍然认为生物科学与社会科学之间的鸿沟将是一种永久的不连续性,立足于认识论并为专家们这一边目标的根本性差异所强化。我们反而视之为一种鲜为人知的进化过程——一种复杂的、迷人的交互作用,文化在其中为生物学指令所生成并塑造,而生物学特征又同时为对文化创新做出反应的遗传进化所改变。
除了少数例外
 ,进化生物学家们都在犹豫要不要将生物学起因与自然选择的概念扩展到文化研究。他们是被一种可称为普罗米修斯—基因假设的东西的流行给抑制住了:遗传进化产生了文化,但只在创造着凭文化进化的能力的意义上是如此;于是一群普罗米修斯基因就从其他基因那里解放了人类的心灵。对于他们这一边而言,又一次除了一些值得注意的例外
,社会科学家们在此观点上达成了一致意见并申明了社会科学的自治权。他们所持的是一种由一条第二关键假设,即人类的灵通统一性演绎而来的社会进化的生物学无量纲观点。该观点坚信,人类文化曾在一个对于遗传进化而言太短而发生不了的时间内进化,而在任何情况下它都大大依赖一个单一的普罗米修斯基因型。
,进化生物学家们都在犹豫要不要将生物学起因与自然选择的概念扩展到文化研究。他们是被一种可称为普罗米修斯—基因假设的东西的流行给抑制住了:遗传进化产生了文化,但只在创造着凭文化进化的能力的意义上是如此;于是一群普罗米修斯基因就从其他基因那里解放了人类的心灵。对于他们这一边而言,又一次除了一些值得注意的例外
,社会科学家们在此观点上达成了一致意见并申明了社会科学的自治权。他们所持的是一种由一条第二关键假设,即人类的灵通统一性演绎而来的社会进化的生物学无量纲观点。该观点坚信,人类文化曾在一个对于遗传进化而言太短而发生不了的时间内进化,而在任何情况下它都大大依赖一个单一的普罗米修斯基因型。
虽有普罗米修斯—基因假设与灵通统一性假设直接的貌似合理性,它们假定的条件还只是特殊的、极端的情况,仍然有待于一种耦合了遗传进化与文化进化的更为广泛的理论的评价。这样一种“比较社会理论”将视人类为一群物种之中的一个,既真实,又可以想象地是可进化的(Wilson,1980a)。自然科学的经验教导我们,当现实世界在可能世界的矩阵中被可视化时,最强的理论就被创造出来。当一组基因—文化特性被遍及许多可想象物种地推断出来并超越了人类变异的任意限制,且被用来界定描述进化过程所需的数学因素时,人类历史的弹道就能够被识别出来。
动物行为学家与社会生物学家大体上赞同了这一立场。他们把人描述为各种各样灵长目物种当中的一个物种,每一个都以特异性的方式适应于特有的环境。为评价人类行为的特有性,他们往往会讲到有着人类特定遗传基础的行为模式,因而讲到规定行为的那些基因。当学习被完全讨论,它大体被视为一种编码外在行为的信息包以病原体侵入宿主的相同方式在两代之间跳跃并拓殖大脑的过程。
至少对于人类而言,这些假定是根本不正确的。行为在基因中并不明确,而心灵不能被视为仅仅是行为特征的复制品。在这本书中,我们提出一种非常不同的视角,以此看来,基因规定了一套生物学过程,我们称之为后成规则,指导着心灵的装配。这种装配是背景依赖的,随后成规则一道以采自文化与物理环境的信息为食。此类信息被锻造成为也就是思考与决定的原材料的认知图式。自发性行为只是心灵动力学的一个产品,而文化是后成规则向着精神活动与行为的团块模式的转译。相对于传统的动物行为学与社会生物学进路,包括先前的基因—文化协同进化进路,我们考虑了心灵的自由区间活动以及由它们创造的文化的多样性。基因确实是,但是是以一种深奥而微妙的方式与文化连接在一起的。
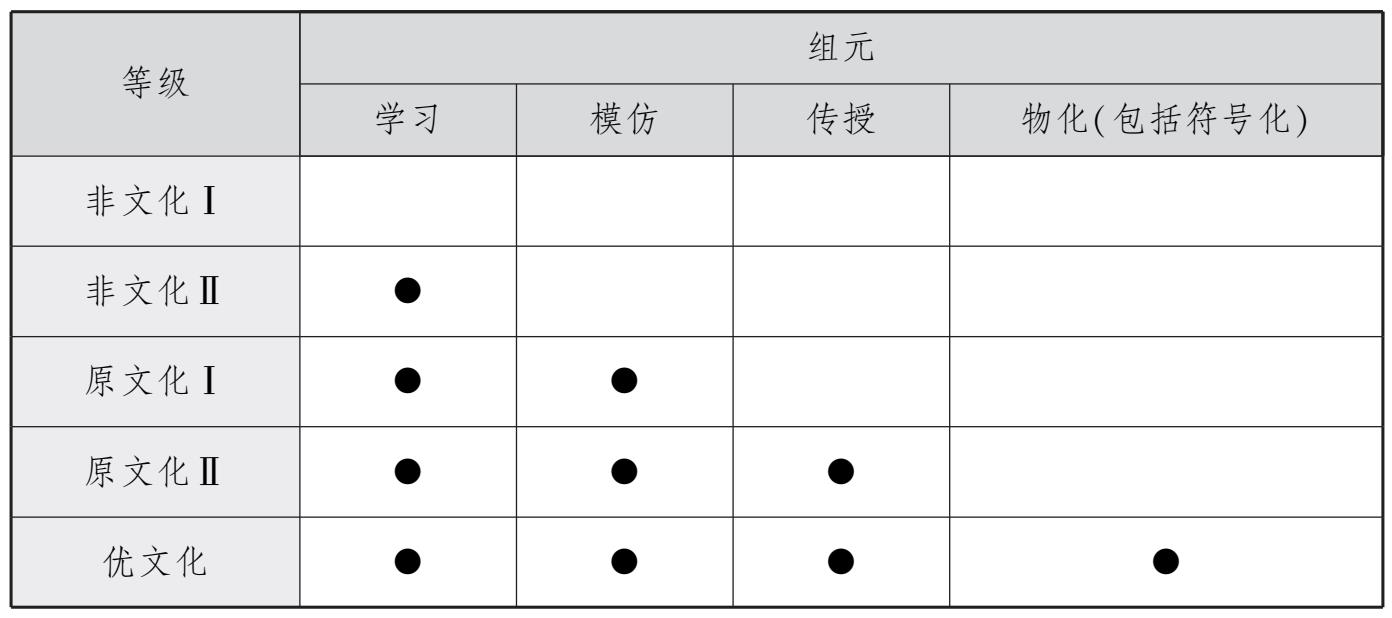
我们将通过一种简单的进化分类来着手启动我们的事业。设想一系列在教与学的能力上存在差异的物种。它们能够被安排到拥有文化进化四种基本组元不同组合的几个集合之中:简单学习、模仿、传授与物化。最后一个术语意指符号以及环境的其他抽象表达的建构,一种我们马上将以更多细节讨论的功能。这些组元中的每一种都可以想象会独立进化于所有其他的之外,不过在动物进化的大多数,如果不是所有的种系发生线中,它们按照我们刚刚以此给出它们的那种顺序出现了:简单学习→模仿→传授→物化。在表1.1中,这些组元的组合被运用来界定5个进化等级,它们粗略地平行于文化行为的逐渐涌现。为我们所知的每个物种(见以下评论:Wilson,1975;Alcock,1979;Beck,1980;与Bonner,1980)都落入这些等级的一个或另一个之中。在作为一个整体的动物界中,从非文化等级(Ⅰ+Ⅱ)到优文化等级(见图1.1),物种的比例在每一步上陡然下降。人独自达到了优文化等级,那就是,最先进的或“真正的”文化状态。
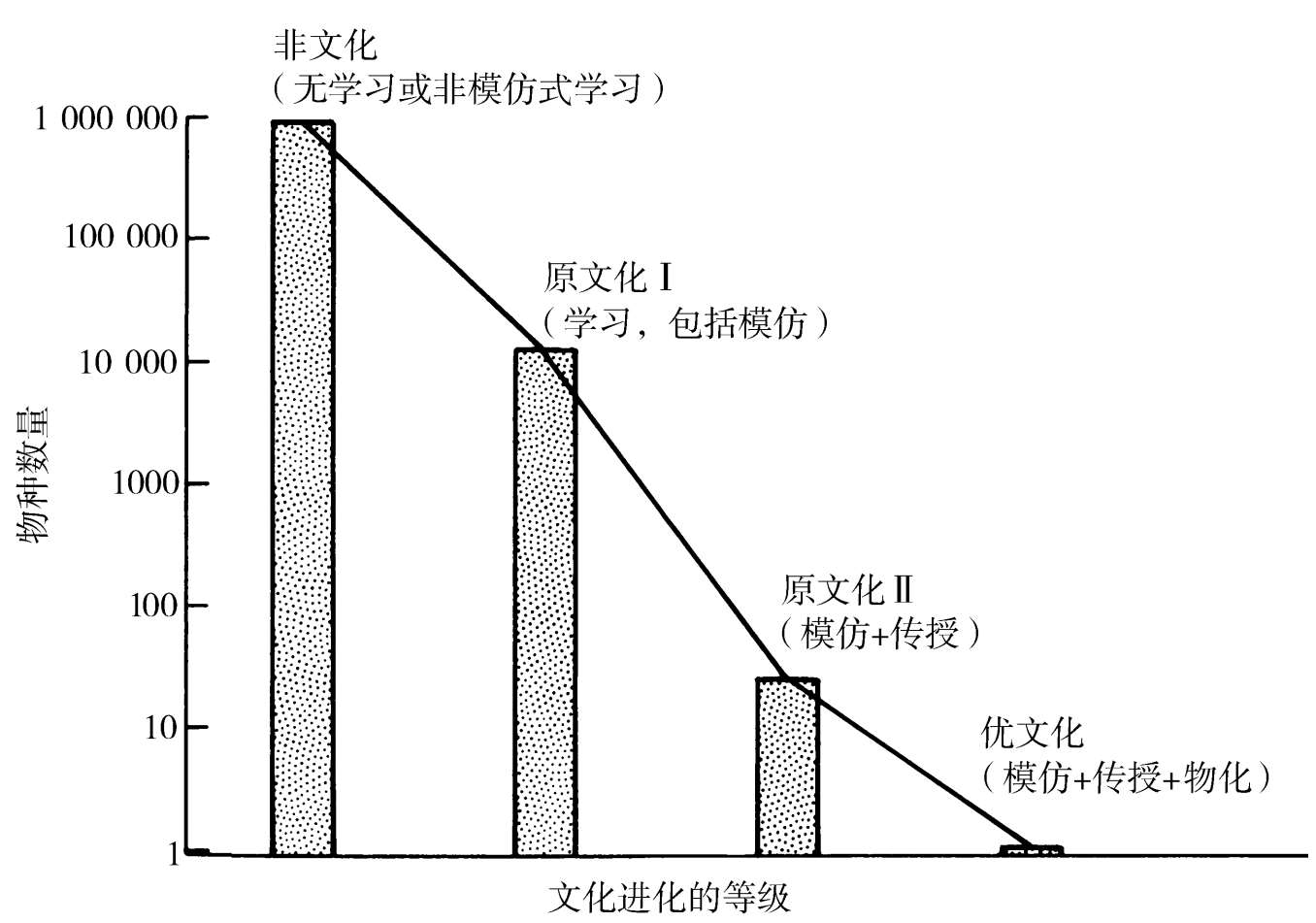
图1.1 文化行为主要等级中现存动物物种的数量。非文化组被视为由所有无脊椎动物以及冷血脊椎动物组成;这组中的1000000个物种是一个数量级的估计(见例如Wilsonetal. ,1978)。原文化Ⅰ组被视为粗略地包括8600种鸟与3200种除原文化Ⅱ组中那些之外的哺乳动物,其中暂定放入7种狼或狗( Canis )、唯一一种非洲野狗( Lycaon )、一种豺狗( Cuon )、一种狮子、全部两种象与全部11种类人猿。人是唯一的优文化物种。
表1.1 界定文化进化等级的学与教的组元。最先进的等级(优文化)可以通过若干进化路径中的这一条或那一条到达,形成一种阶梯式的组元积累。
我们在广泛的意义上定义文化,包括精神结构与行为的总和,也包括加工品的建造与使用,通过社会学习从一代传递到下一代。尽管认识到符号在人类文化中的杰出性,但我们并不赞同格尔茨(1966;1973∶89)、施奈德(Schneider)(1980)与其他一些社会科学家视它们为唯一的文化诊断法。这种语义的限制武断地排除了实质性的一类模仿行为,其中有些不知不觉地进入符号化的等级。它低估了认知过程的复杂性,其中许多不能被分析性地归入通常给予符号化的意义。最重要的是,这种狭义的定义将关键词——文化——排除在了对动物以及祖先人类的理论分析之外。
人类达到了优文化,部分地是通过实施于年轻人社会化期间的教育来实现的。有意的指导程序,至少由非正式的提问与回答组成,为所有的人类社会所使用(Williams,1972a,b;Davidson,1977;Hansen,1979;Patricia Draper,个人通信)。成年人有强烈的倾向去提供复杂的指导,而年轻人倾向于追随他们。如同沃丁顿(Waddington)(1960)曾适当地表达这种关系,人是负载权威性的物种。
事实是,教育并非为人类所独有。广义上蜜蜂跳摆尾舞时就可以说是在指导巢友。直线的行进,即8字舞模式中心横段的方向与持续时间,象征着新发现的蜜源或某个其他目标的定位。其他蜂儿自动跟随这舞蹈,记忆这些编码的指导,然后按此信息行动起来寻找目标。黑猩猩与其他灵长目的母亲走得更远,运用信号与体罚来指导幼崽玩耍与模仿的动作。例如,在坦桑尼亚的贡贝河国家公园,当婴儿黑猩猩爬得太高时,它们的母亲就敲敲树干,让它们立刻下来(Goodall,1965)。如同邦纳(Bonner)(1980)所指出,教育被多样化为一个穿越少数实践它的动物物种的连续统,范围从表现为带有明显诱导模仿功能的行动,到模仿与探索行为的一步一步强化。但即使是最精致的动物教育模式,也远不及人类社会训练出来的密集而又复杂的指导程序。
简言之,人类在优文化过程的量级上定量地不同于动物。除此之外还有一种独一无二的活动,充分地将人类从最高级的原文化动物物种中分离出来,并使其成为唯一已知的优文化物种。这就是我们称之为物化的过程。人类心灵的操作合并了(1)概念的生成与(2)连续变动着的世界再分类。昆虫、冷血脊椎动物与其他大脑相对较小的动物,在周围感觉细胞与较低关联中心的水平上过滤掉了大多数的信号,进而主要对剩余信号当中一套非常受限的“符号刺激”(sign stimuli)做出反应。相比之下,人类心灵吸收数量多得多的混乱定时的刺激,其中大多缺乏直接的相关性,而从它们之中建构起一种内在的实在性。逐渐变异的构造被复杂但具体的一套操作碎解成不同的类别,其正在开始接受认知心理学家们的深入研究(Posner,1973;Rosch et al. ,1976;Getty et al. ,1979)。诸类别经常成对出现——例如,群内/群外、孩子/成人、神圣的/亵渎的,同时它们的边界也为仪式与禁忌所增强。然后全新的产品,“心智品”(mentifacts)(Huxley,1958)就被创造出来,包括那些情感强烈但只是被心灵的理性部分无力理解的心理过程。如此,诸神、精灵与图腾可被诠释为使心灵的群体绑定活动神圣化的外在表现(Lenski and Lenski,1970;Rappaport,1971)。隐喻被创造来联系那些被更为直接地觉察到的物理现象与更不易被把握到的那些。例如,一个人想象出思想的猛冲越过页面,或代表着部落世袭统一性的鹰,或恰当微分, dx / dt ,表示着一个任意选定的实体中的变化。
物化的可行手段是符号化,其即刻服务于辅助记忆、触发情感、分类环境,以及传送信息与感受给其他人(Needham,1979)。人类语言大体上是符号的操纵,用来传达心灵的物化结构。黑猩猩语言训练实验的一个结果,是纯粹语言机制的部分贬值。依靠符号语言的方式,这些动物能够被教学会超过100个单词,以及传达情感或提出请求与命令的三个或更多单词的初等类句式组合(Savage-Rumbaugh and Rumbaugh,1978,以及Terrace et al. ,1979)。它们明显缺乏的是物化其经验为新的概念,其可依靠无论什么的任何方式,包括单一的符号或句子传送给其他人,这种人类能力与纯粹的驱动力。尽管语法规则与语音是必不可少的传送手段,抽象化与符号化似乎才是优文化进化中的初步成就。
人类凌驾于黑猩猩与其他高级灵长目之上的先进性,获得于一段异常快速的进化爆发期。在一段近乎超过300万年的时期内,从祖先南方古猿( Australo pithecus )以物质为基础的文化开端时起,到现代智人( Homo sapiens )的旧石器时代晚期,大脑尺寸已是原来的3倍。南方古猿的颅容量为400—500立方厘米,相当于黑猩猩和大猩猩的水平。200万年后,直立人( Homo erectus )达到了1000立方厘米的容积。又一个100万年,看得见尼安德特人(Neanderthal man)中一个达到1400—1700立方厘米的增长,现代智人中则是900—2000立方厘米。现代智人的年轻者也要经历一个长得多的社会化时期,一个明显起源于人类新皮层扩大之时的阶段。黑猩猩、红毛猩猩和大猩猩的幼年期有7到8年。在现代人中,它大约是14年,并伴随以认知与生理学发育一种相对缓慢的、设定程序的展开。此外,这种发育还包含了导致高级认知、思考与学习水平的全新要素(见图1.2)。
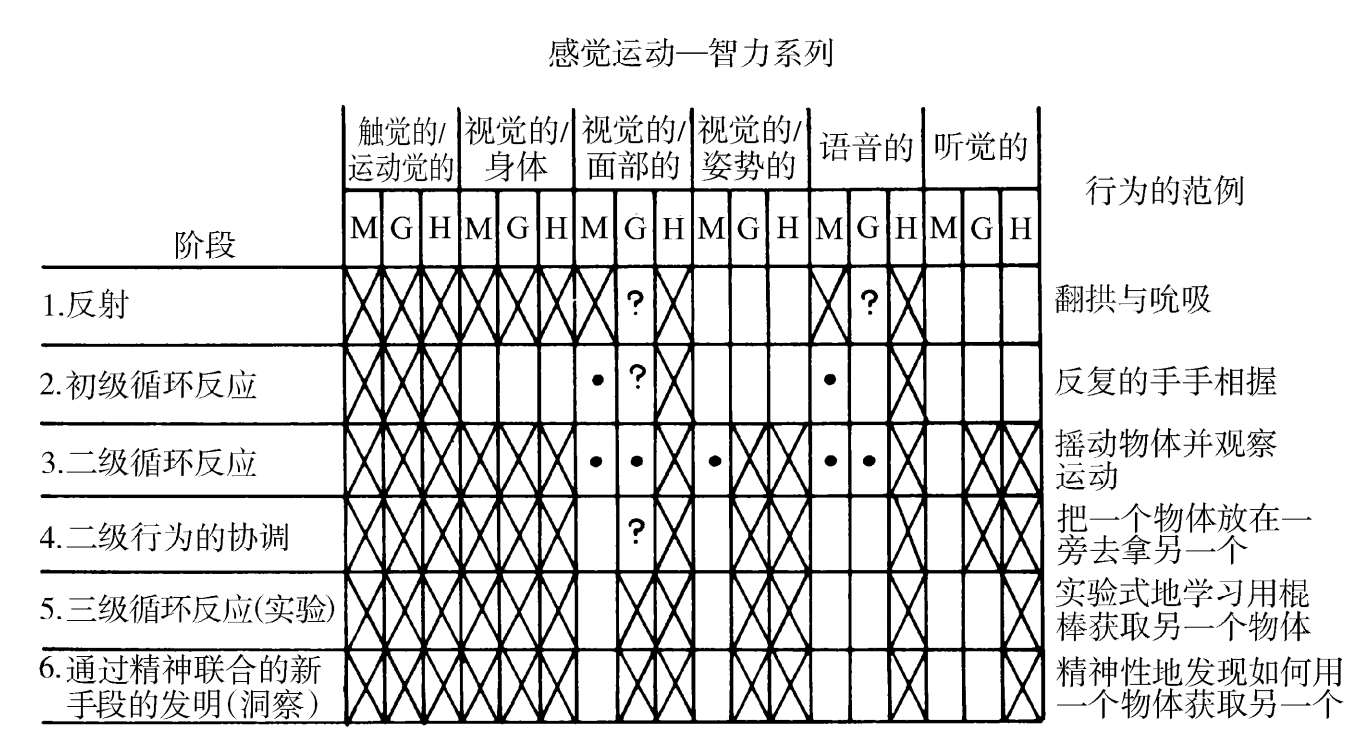
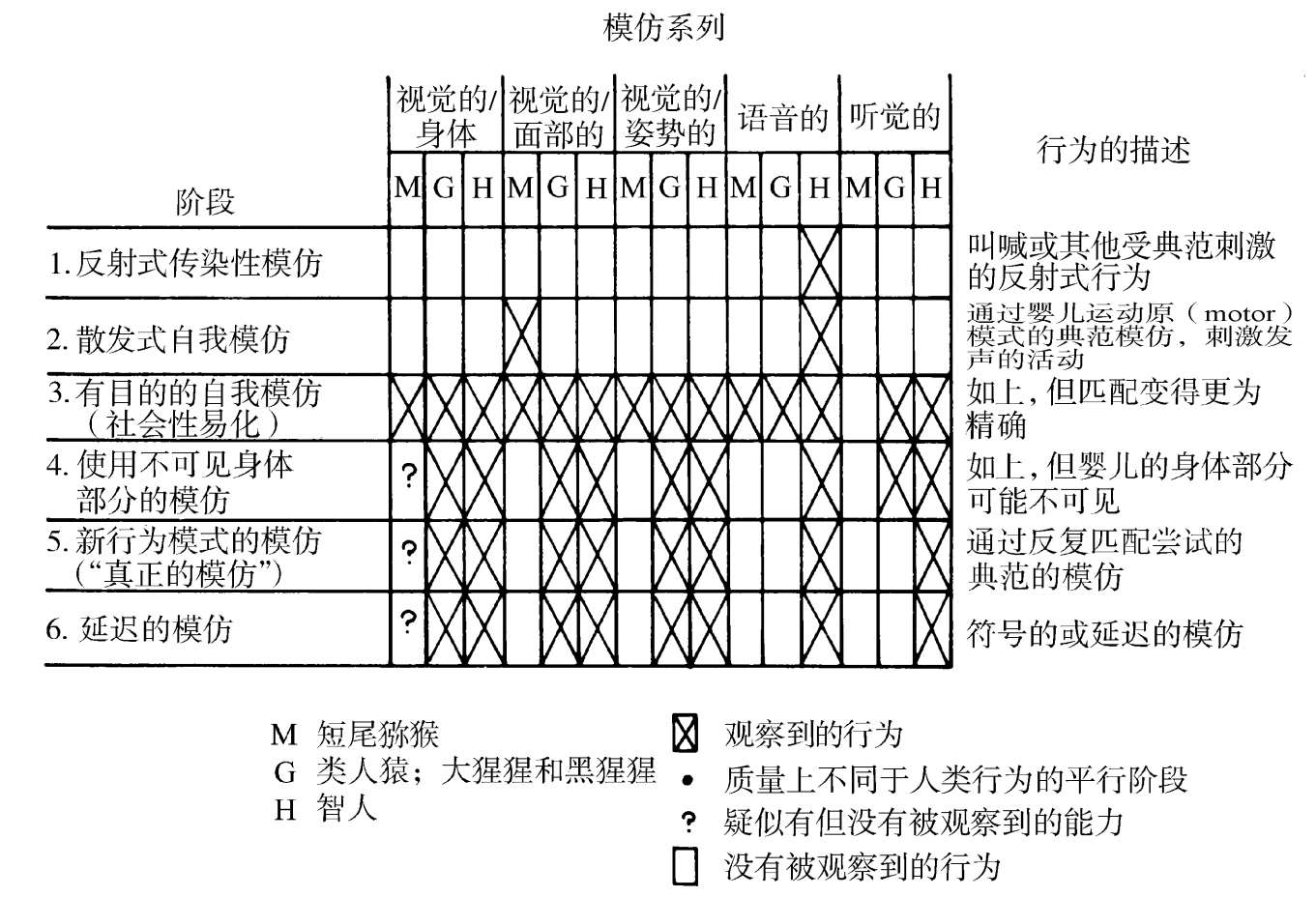
图1.2 年幼的猴、类人猿与人类的发育程序的比较。所用分类采自皮亚杰(Piaget)的那种,并续以感觉运动智力与模仿能力发育的系列阶段。(基于Chevalier-Skolnikoff,1977)
简言之,凭借单一的智人物种达到优文化,在不止一个方面都是一个独一无二的事件。它是通过生命史上一次前所未有的神经解剖学与行为的进化加速来实现的。一个人可以用几乎是物理学的术语来可视化这一过程:凭借此物种的优文化阈值的穿越,跟随——或许不可避免地——以一种持续的自催化反应,遗传进化与文化进化在其中驱动彼此向前。
为了更严密地考察这一显著的过程,有必要考虑遗传进化与文化进化得以贯穿个体发育的程序而互动的方式。如图1.3所示,在一个社会物种中有三种程序是可能的。想象此刻有一组可传递的行为、心智品与加工品(artifacts),我们提议称其为 文化根 (culturgens)[来自拉丁语 cultur ( a ),文化,加上拉丁语 gen ( o ),生产;而被我们发音为“kul′tur jens”]。此单位为考古学中所采用的加工品类型(artifact type)的对等物(Clarke,1978),在可变程度上还类似于布卢姆(Blum)(1963)的记忆类型(mnemotype)、赫胥黎(Huxley)(1962)与卡瓦利-斯福尔扎(1971)的理念(idea)、默里(H.A.Murray)(于Hoagland,1964)的理因(idene)、斯旺森(Swanson)(1973)的社会基因(sociogene)、克洛克(Cloak)(1975)的指令(instruction)、博伊德与里彻森(1976)的文化类型(culture type)、道金斯(Dawkins)(1976a)的模因(meme)与希尔(Hill)(1978)的概念(concept)。附录1.1中给出了使用这个新词的正当理由,以及它更为精确的定义。图1.3中简化表示的文化根对于教学双方而言同样可及。它们要通过一系列 后成规则 来处理,其为遗传决定的程序,指导着心灵的装配,包括对通过周围感觉过滤器的刺激的筛查、神经元间的细胞组织化过程,以及更深的定向认知过程。这些规则构成了基因置诸发育的约束(因而有“后成”的说法),并且它们影响着使用一个相对于另一个的文化根的概率。这种概率分布自身被适当地称为 使用偏向曲线 (usage bias curves),或者更简单地,就叫 偏向曲线 。
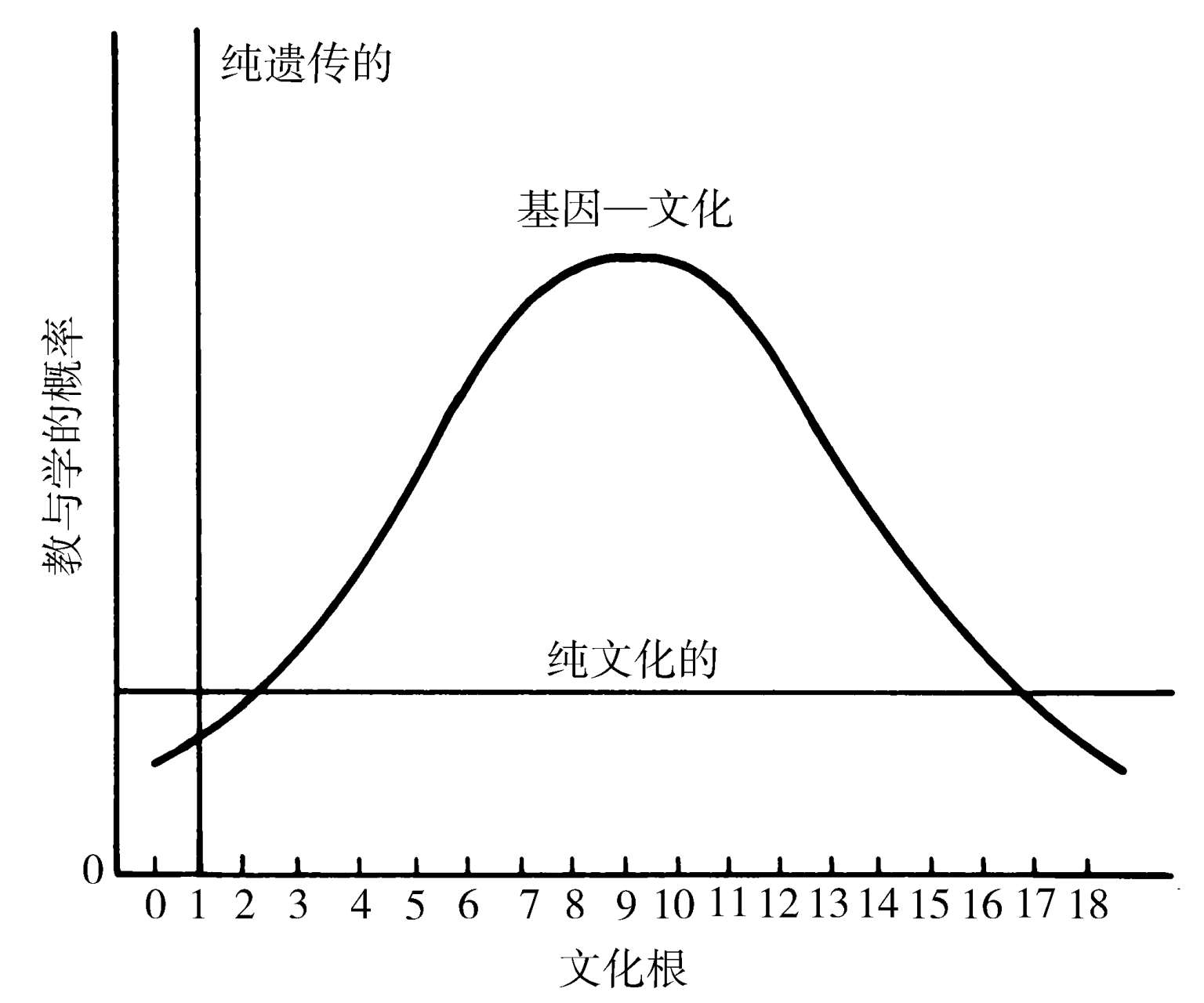
图1.3 社会物种中三种可想象的信息传递程序。根据后成规则(遗传决定的细胞周围筛查与定向认知的过程)的特性,文化根传递的概率将被限制在一种单一的选项(纯遗传传递)、所有选项等概率(纯文化传递)或并非等概率的多个选项(基因—文化传递)。这些曲线是为给定有关特定文化根类别的基因型的个体生物所拥有的后成规则的产物。(修改自Lumsden and Wilson,1980a)
设想一物种内给定基因型的幼稚种群或幼稚个体面对着一套文化根。这些要素可能是一种食材搭配、一套木工工具、各种各样可被采用或抛弃的替代性婚姻习俗,或任何可比较的一组选项。如果个体社会成员的发育是以每次有相同的文化根被选择这样一种方式被遗传地约束,这种传递就被说成是 纯遗传 传递 。后成场,即是所有发育选项的时程,是一条从出生到成熟伸展着的单一的狭窄渠道。在非文化物种中,受到类似约束的行为选项,被动物行为学家们称为固定行为型(fixed-action patterns)。取决于物种与行为类别,此类反应能够全部地在中枢神经系统中被编程,而不需要学习无论什么。或者,如果有学习发生,它会被如此紧密地指导,以至于只有一种基因型特定的行为才能实现。干涉发育过程将导致发育不全版本的行为或完全没有行为,换成某种替代性的充分发育模式(Alcock,1979)。如此,许多鸟类物种的幼崽必须学习本物种特有的歌唱,因为它们没有能力学习任何别的。纯遗传传递到目前为止已是生物学家们的主要兴趣,尤其是神经生物学家与动物行为学家们,因为它典型地与“本能”密切相关,是文化的相对反面。但要注意,一种高度社会性的物种也可以想象地是会进化的,其将拥有一种高级的语言与文化,所有这些都是学来的,并且也能够传递仅仅一套行为以至一种文化。换句话说,纯遗传文化是一种逻辑可能性!在一种被完全约束的物种中,因为后成规则中的一个变化,无论它被编程为固定行为型还是充分渠道式学习,从一种被偏爱的文化根到另一种的转换,就是纯遗传进化。
在相对的极端上,图1.3中也有所表明,所有的可用文化根都将被同样可能地利用。后成规则进化如此,以至在个体发育中移除了所有形式的偏向性,其原本可能产生于周围感觉筛查、神经元间细胞组织化或更深认知过程中的内在倾向。这种 纯文化传递 正是许多社会科学家在他们诠释人类进化时心中所存有的东西:
人性在社会—文化上是可变的。换句话说,并没有在生物学固定基底决定社会—文化形成可变性这种意义上的人类本性。只有在限定与许可人的社会—文化形成这种人类学变量(例如,本能结构的世界开放性与可塑性)的意义上的人类本性(Berger and Luckmann,1966∶46—47)。
这一假说似乎是一种盛行的看法。在一项关于24本导论性社会学教科书的分析中,彼得沙克(Petryszak)(1979)曾发现下述假设是基础性的:“关于被相信内在于人类物种的生物学因素的任何考虑,在理解人类行为与社会的本性方面都是完全不相干的……人类文化是单独地由社会的观念性与技术性方面所构成的,而不包括关于有一个生物学基础的任何考虑……人的学习以及对于社会化过程与他人意见易感的能力,只是由于所有本能的缺席。”在这样一种整个不受约束的物种中,文化根偏爱性的转换就会是纯文化的进化。乍一想,这样的一种安排,可能似乎构成了一种相对简单、普遍又稳定的状态,既然社会行为从基因那里“解放”了出来。但相反的情况更有可能是事实。文化根中偏爱性的生理学平等化,需要一种编码多个复杂的微调机制的基因型。只有在依靠连续遗传进化的精确控制下,纯文化传递才有可能维持。
介于其间的情况是
基因
—
文化传递
,定义为在其中多于一种文化根可使用而至少两种文化根因内在后成规则而在采用的可能性上有所不同的一种传递。
基因
—
文化协同进化
被相应地定义为起因于基因频率变动的后成规则之中,或者起因于后成规则的文化根频率之中,或者共同存在于二者之中的任何变化。如我们将要证明的,给出足够的时间,这两种变动都会不可避免地发生,并且它们也会彼此施加以一种互惠式的影响。关于这一交互作用正式的概念化与分析,可被称为基因—文化协同进化理论,或者更简洁地称之为:
基因
—
文化论
。
在先验的基础上,基因—文化传递似乎对于所有类别的文化根而言,在人类物种以及在所有其他可想象的、身上有文化能力在进化的物种身上,都是最有可能的传承模式。由于如下原因,纯文化传承是一种不大可能的遗传进化备选结果。
(1)在人类感觉形式(视、听、味、嗅、触、潮湿感、热感)的每一种之中,敏感性都在大量可接收刺激的范围内发生着程度上的变化。在视与听的案例中,狭窄的上限与下限括住能被区分或甚至被知觉的波动频率。对于一个进化着的生物学系统而言,依靠转导细胞与神经元链,要想获得充分的质量控制以接近一致的敏感性,与此同时取消频谱上的限制,将是困难的或不可能的。同样的考虑也适用于中间神经元之间的编码装置对模式与复杂性的读取。简言之,中枢神经系统先天地就没有准备要平等地知觉与分类来自文化根全范围的初级刺激。一定会有一些偏向性, 自始 内置于它们的筛查与编码设备之中。
(2)智人,以及理论上可以想象的、极其依赖文化传承的任何其他物种,都不可能从一种无差别的行为发育来开始它的进化。如同在所有的动物物种中那样,直系的祖先种群必定依赖了自动的周围筛查与模式编码,以及有准备的学习形式,诸如产生于一种适应性的、对于某些刺激的偏爱的印记与抑制。
(3)即使一个物种能够以某种方式启动于一致的后成规则,该策略一般也将是不稳定的,会在基因—文化协同进化的进程中导致不一致的后成规则的再次出现。
设想一个二倍体生物的种群,带有个体基因型 G i G j ,这里每个成对的 i 和 j 代表每个位点上的等位基因。在一个文化物种中,一个生物体的遗传适合度 w 不仅由它的基因型 G i G j 决定,而且由它的文化遗产决定,表现为它的被同化文化根的集合,c o 遗传适合度 w 服从于缘于学习、创新与其他时间依赖的更改文化根内容的文化过程的变化:
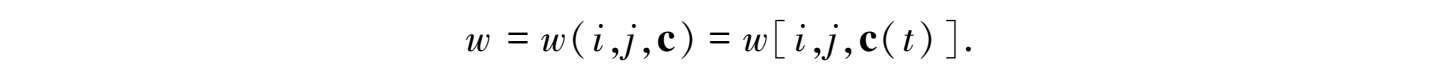
如图1.4所示,当关于文化根集合从c 0 到具有同等或较高相对适合度的一个集合c k 的变化的概率每单位时间 v 0 k ( t ),大于关于到具有较低相对适合度的某个其他集合c m 的一个变化的概率每单位时间 v 0 m ( t ),则遗传适合度会被提高。后成规则提供这一能力,以向着相对有利的集合如c k 的转变比向着相对有害的集合诸如c m 的转变具有更高频率的发生这样一种方式,塑造着生物体的文化根使用模式。
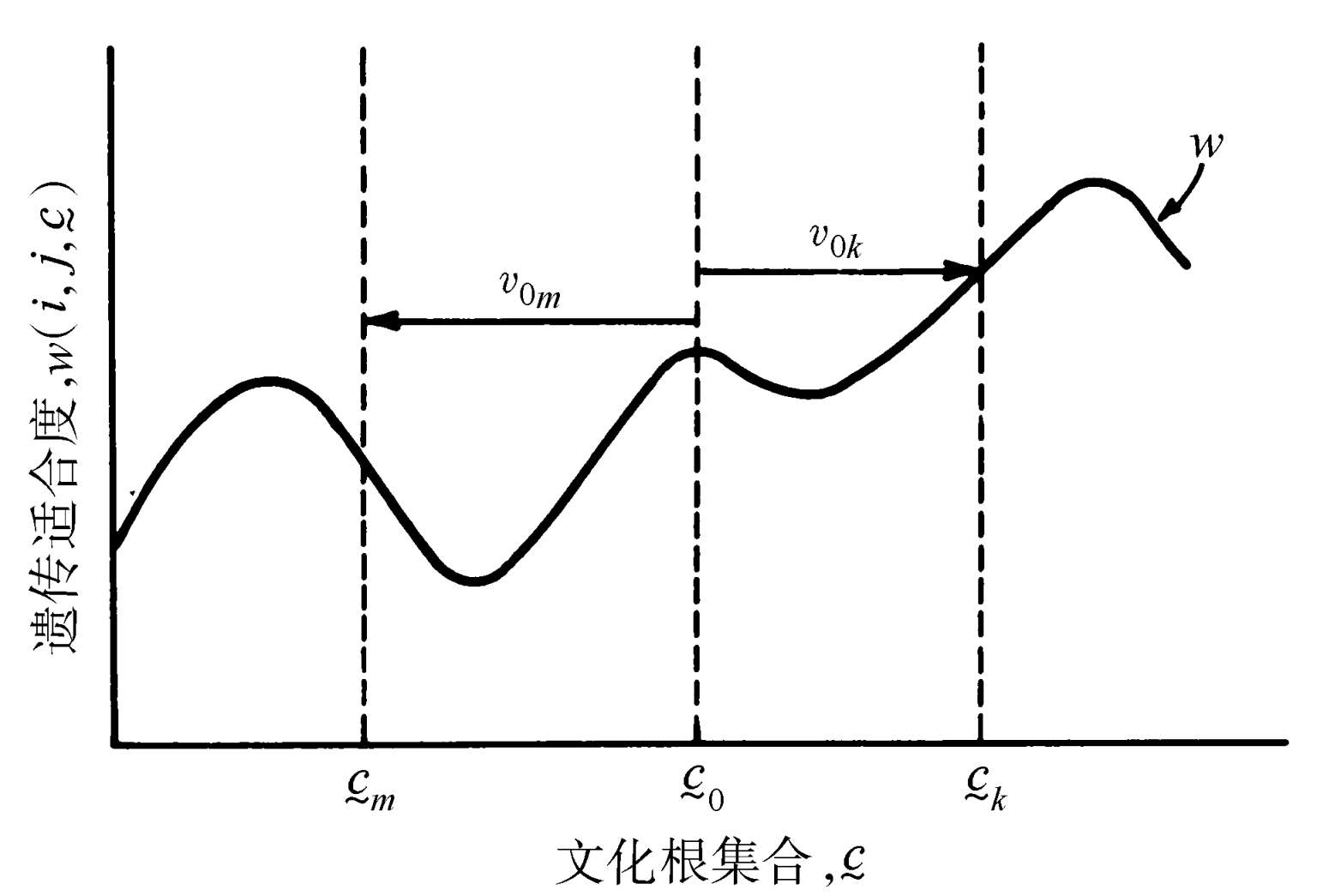
图1.4 后成规则的进化。自然选择偏爱一致地使从文化根集合c 0 到文化根集合c k 的变动概率v 0 k 大于从c 0 到c m 的变动概率 v 0 m 的生物,这里由c 0 所赋予的遗传适合度大于由c m 所赋予的,但小于或等于由c k 所赋予的。
现在设想一个白板生物体种群,其改变着它们的文化根集合,并变动着加之于由不涉及对遗传适合度的影响的特定文化根所发动的行为之上的控制程度。换句话说,该发育场是平的。该种群暴露于这样一种环境,既包含适应性的,又包含有害的文化根,但它并不能区别它们。与此同时,它对能够一时兴起设置 v 0 m > v 0 k 的文化编程又是开放的。经过数代时间,该种群在遗传突变的入侵面前,其会编程后成规则偏向于向着相对适应的集合同化的个体,是不稳定的。然后,后成规则会倾向于将认知发育引向相对于其他的某些文化根。为了使它在比喻上更加生动,我们非正式地称这种关系为“皮带原理”:遗传自然选择以这样一种方式操作,就像是控制文化于一条皮带绳之上。
这皮带象征着遗传规定的、使用承载某些贡献遗传适合度的关键特征的文化根的倾向性。这些倾向性有别于必定导致特定文化根同化(而由此完全不允许皮带)的硬连线算法(hardwired algorithms)。在文化物种中,几种其他的因素将无论如何都会合起来支持一种皮带的延长。用纯生理学的话说,从一个能够识别并分类文化根适应性特征的神经子系统那里增加的利益,随着其精确度的增加,将最终为创建系统所需的个体发育路径成本,以及维持它于一个功能性状态的新陈代谢成本所抵消(见图1.5)。分辨能力的这种极限,在文化根同化能力上产生出额外的灵活性,也增加着皮带的长度。在优文化与一些原文化生物的案例中,我们预期文化根集合将不只恰恰是一个同化文化信息的被动容器。它也是一个新的文化根在此能够被发明出来的场所。对于创新着的文化生物而言,一个文化根的适应值,部分地取决于生物体“播种”拥有适应性特征并促进其同化的新事物的潜力。即使当新颖的文化根承担高风险之时,在先前曾受遗传自然选择偏爱的后成规则之下,它们也能携带一种相对高的同化概率。在风险成为现实之前,此类文化根也许会,也许不会结出短期的果实。
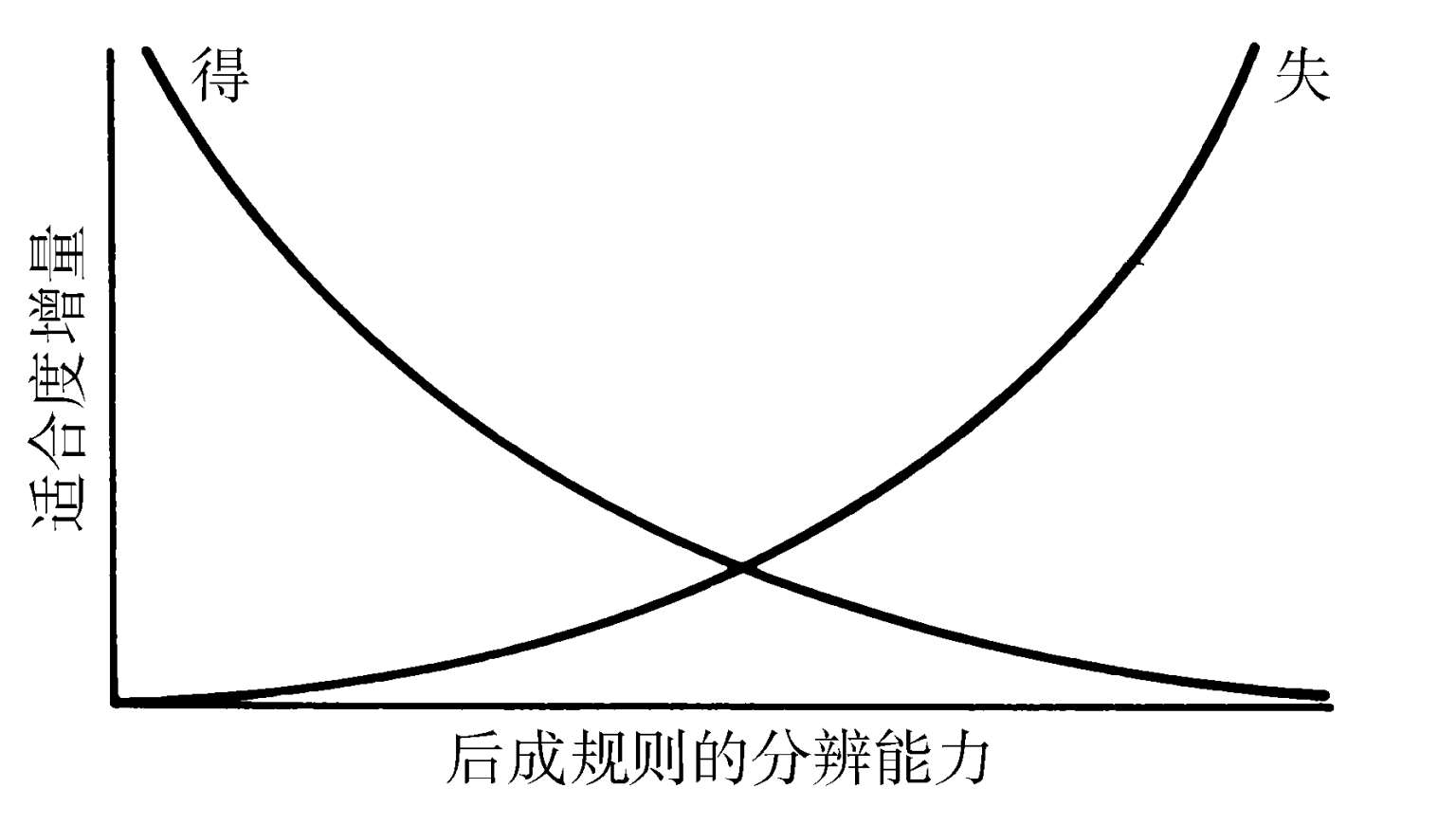
图1.5 为一种遗传适合度净收益的最终减少所约束的后成规则分辨能力。这种减少既缘于从提高分辨率中获利的增量下降(图中标为“得”),也缘于分辨率提高所需的更精确生理学机制的上涨的成本(标为“失”)。在这里描绘的假想案例中,当曲线交叉时,或说在一种中间分辨能力的水平上,则最适度被达成。
从白板状态中,转变概率可能要么为适当的新基因型的并入,要么为新文化根的发现所扰动。前者通过改变影响文化根使用概率的后成规则,变动着对先前存在文化根的遗传适合度的影响,同时后者也变动着先前存在基因型的适合度。重点在于,两种事件都会将种群从白板状态中移出。
我们甚至能估计出文化根创新将有此扰动性影响之前的等待时间。此类信息使得以一种初步的模式评价那些贡献性因素的相对重要性成为可能。在文化根创新的案例中,这里文化根与另一个之间的差别随机可变且文化根被以恒速创造,一个适合度变动着的文化根的引入时间为
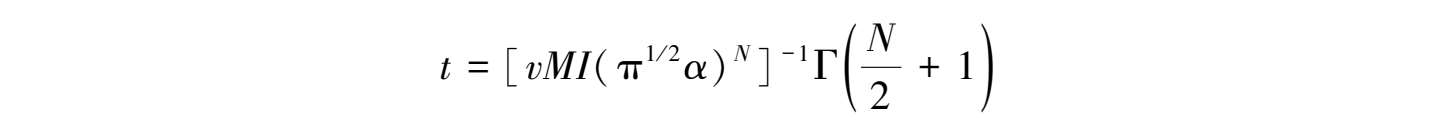
且这里 v 为单位时间内每人文化根中的文化根创新率, I 为种群人数, M 为可能得到的适合度变动着的文化根数量, N 为一个文化根得以区别于他者的品质的数量, α 为关于区别个别文化根的难度的一种量度,Γ代表伽玛函数。如图1.6中的曲线所示,这些系统对于区分文化根的能力上的变化似乎是最敏感的。随着文化根空间维数( N )的增加,以及在不同维度上认知简易度的提高(通过 α 的减小),适合度变动着的文化根被添加的频率下降得非常陡峭。相同的原理,也适用于作为一种极端的、危害全社会福利的适合度变动着的创新的“死文化根”的同化。文化根区分能力的这种提高,会放慢从白板状态中的离开,但不能完全避免它。(关于等待时间估计公式的一种推导,见附录1.2。)
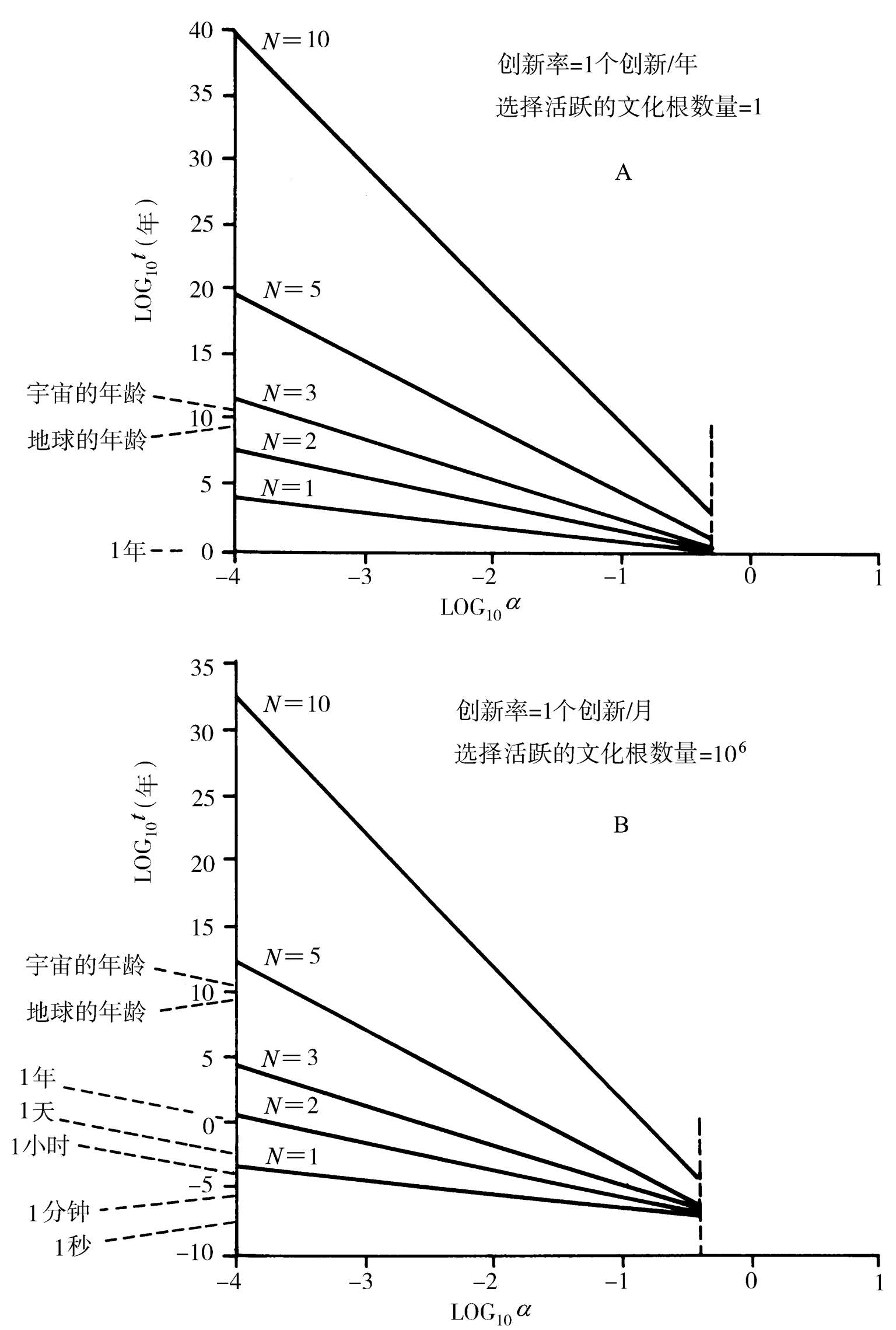
图1.6 一个社会从一种白板状态中离开的等待时间,作为用来区分文化根的属性的数量 N 与认知系统的分辨力 α 的一个函数。显示于图表的创新率是整个种群的那些,换句话说,是 vI 。
如前述讨论所示,人类物种是置身于基因—文化协同进化之中的吗?在我们看来,对于这样一个结论,最低限度上,四种证据将是必要的。第一,必须表明,不一致的后成规则存在,它们是普通的,如果不是普遍的,并且它们能够以这样一种方式被分析来检验基因—文化协同进化理论的细节。我们将在第2章和第3章中证明,这第一个要求是充分满足的。
第二个要求是,人类种群内部后成规则的遗传变异必须存在。因为后成规则研究仍然处于早期阶段,其变异的范围与原因很少被明确地考虑了——并且当然不是在基因—文化论的语境中。但是谱系分析与异卵及同卵双胞胎的标准比较,在某些情况下为发育的纵向研究所增强,凭借这种手段,包括要么构成后成规则要么与其共享组元的一些,被研究的几乎每一类别的认知与行为中,都产生了遗传变异的证据。这些类别包括色觉、听敏度、气味与味道区分、数字能力、词语流畅性、空间能力、记忆、语言习得的定时性、拼写、造句、知觉技能、精神运动技能、外向性/内向性、同性恋、酗酒的倾向性、初次性活动的年龄、皮亚杰式发育阶段的定时性、一些恐惧症、某些形式的神经症和精神病,包括狂躁—抑郁行为和精神分裂症,等等(Heston and Shields,1968;McClearn and DeFries,1973;Ehrman and Parsons,1976;Farley,1976;Loehlin and Nichols,1976;Martin et al.,1977; Bohman, 1978; R.S.Wilson, 1978; Ashton et al. ,1979;Comings,1979;Rainer,1979;Vandenberg and Wilson,1979)。
基因—文化协同进化成立的第三个条件,是人类种群内部文化实践与遗传适合度之间一种关联性的证实。事实上,在一大批行为类别之中,这样一种关系已经被记录下来了,且经常牵涉到文化根之间的细微差别。例如,在文身与其他身体标记模式中的某些实践,如同在割礼、对月经及产后血的处理,以及日常饮食中,已知都传播着病毒与其他极大影响死亡率、出生率甚至性别比例的传染源(Blumberg and Hesser,1975;Gajdusek,1977;Drew et al. ,1978)。关于性行为、婚姻习俗、早期母婴依恋、有差别杀婴、攻击的形式化技术以及经济组织等对遗传适合度的直接影响,记录也存在(Daly and Wilson,1978;Chagnon and Irons,1979;D.G.Freedman,1979;Kennell and Klaus,1979)。然而,此类实践的长期效应还没有得到测量。
相对细节化的关联性,在烹饪案例中的文化与遗传适合度之间建立起来了。人类不能遗传地生物合成赖氨酸这种氨基酸,其必须从一种平衡的饮食中获取。玉米,新世界唯一的栽培谷物,拥有大量的赖氨酸,但2 /3的化合物被锁定在胚乳和胚芽不可消化的谷蛋白片段中。释放这些被隔离部分最简单的方法是碱法蒸煮(alkali cooking)。此技术的数个变种被发明出来并通过文化的扩散而传播开来。在卡茨(Katz)与助手们评论过的北美洲、中美洲及南美洲的51个社群当中(1974),玉米耕种的强度、碱法蒸煮的使用、人口密度与社会组织的复杂性之间存在一种强正相关。考古学数据与当代社会的观察显示出,当高度的依赖性被加在作为一种主食的玉米之上,人口密度就会增加。但除非被加工以释放赖氨酸,玉米并不是一种有营养的食物。看起来,许多采用碱法蒸煮的新世界社会,都不大可能会直接将此道工序理解为其生化短缺的解决乃至促进人口增长与社会进化之必需。
一个不同种类的基因—文化关系体现在蚕豆,地中海地区最流行也最容易耕种的作物之一的食用上。基因G6PD-是性连锁隐性的,当没有等位基因G6PD + 的陪伴时,会导致一种红细胞酶,即葡萄糖-6-磷酸脱氢酶的缺乏。在地中海人口当中,G6PD-的发生率位于0.05到0.30之间。它的相对高频率一般被归因为它带来的更高抗疟疾性。然而,由该基因导致的纯生化缺陷会因蚕豆的食用而恶化,引起疾病与时而的死亡。似乎存在一种G6PD-/ G6PD + 的多态性,随着G6PD-的发生率几乎肯定地为蚕豆的食用以及由此引发的蚕豆病所降低。不足为奇的是,在贯穿大部分有记载历史的地中海社会中,蚕豆已是烹饪特殊仪式、选择性禁忌与民间传说的相关物件。尚少有证据表明,受到影响的人们曾在其信仰与现在被认为是蚕豆病的真实本性和原因的东西之间,建立起任何直接的、理性的关联(Katz,1980)。
关于基因—文化协同进化的第四个,也是最后的必要条件,是直接关联基因与认知发育的分子以及细胞机制的存在。动物物种中感觉接收与行为的中间纽带,已被广泛地记录下来(McClearn and DeFries,1973;Ehrman and Parsons,1976;Hall and Greenspan,1979)。关于人类认知中这样一种连锁性的证据是碎片化的,同时一些基本步骤也已为人所知。即使是最复杂的后成规则,也能为影响分子与细胞机制的遗传变化所更改。例如,在周围感觉筛查的层次上,改变色觉与对苯硫脲敏感性的单一突变,就是以初级感受器细胞的分子结构变化为基础的。同样初等的遗传控制,也会影响到觉察某些气味诸如十五酸内酯的能力,并且可能构成了在知觉类麝香物质环十五内酯方面的性别差异的基础。
在下一个组织层次上,20种或更多种已知的神经递质,包括一元胺、血清素、去甲肾上腺素与多巴胺,有一些会对心情、专注度、睡眠习惯与社会行为施加重大影响。它们运作在特定的感受器位置,而它们的影响能够为抑制其特定氧化酶或以其他方式改善突触持久性的物质的使用所增强。在任何有意义的程度上改变行为有效性神经递质产生或其感受器位置特性的突变,都有可能同样改变了后成规则。例如,最严重的临床上可定义的精神分裂症形式,就是明显地处在部分的遗传控制之下(但见Taylor and Abrams的保留意见,1977)。它们与多巴胺感受器的较高密度或多巴胺的较高浓度有关(Greenberg,1978)。类似的可能性,也存在于促黑素细胞激素,一种影响记忆与焦虑水平的肽,以及调解疼痛感受、焦虑与压抑的内啡肽的调节过程中(Arehart-Treichel,1978)。神经生物学家与生理心理学家们是乐观的,相信为人的行为现象负责的那些细胞机制最终将被理解(见例如Boddy,1978,以及Schmitt and Worden,1979)。法利(Farley)(1976)与其他生物精神病学家们走了很远,以至总结出,神经症与精神病,是由经常恰恰超出了正常的遗传调解活动范围的心理学事件所引起的精神状况。充分的扰动,可能产生于异常基因型、早期发育事故、环境压力或这些过程之间的交互作用。如此,行为中的根本性变化,有时可能会跟随影响着关键细胞活动的相对少量的遗传变化而来。
在一个还要更高的、细胞结构的组织层次之上,大鼠、恒河猴与人类大脑的愉快中枢现在已知并非由局部的细胞群集构成,而是由从边缘区域扩展进入额皮质的整个纤维束构成的。研究显示出,此“大脑奖励系统”中的活动有差别地受到一元胺神经递质的影响(Routtenberg,1978;Stevens,1979)。我们认为,该系统的一个遗传变化将会改变认知发育的方向,如此也会构成后成规则的一次修改。在最高的层次上,也不乏用来解释意识的神经元基础以及解释心灵的模型。确实,这在当前是理论神经生物学最活跃的领域之一,而其范围被示例于普里布拉姆(Pribram)(1971)、布塞(Buser)与鲁热尔-布塞(Rougeul-Buser)(1978)、科尔比(Colby)(1978)、埃德尔曼(Edelman)与芒卡斯尔(Mountcastle)(1978)、格罗斯伯格(Grossberg)(1978)、勒德雷尔(Roederer)(1978)、瓦塞尔曼(Wassermann)(1978),以及西蒙(Simon)(1979)的著作之中。诸模型中的一些格外微妙,并且创造性地动用了大量关于大脑结构及作用的实验数据。身—心关联在细节上仍然有待于解谜,同时其可行性已在基于神经生物学信息的具体重构中建立了起来。
简要地总结这一点,我们讨论了,自其进入最先进的、“优文化的”社会组织等级,基因—文化传递就是人类物种的一个几乎不可避免的特性,因为在任何文化物种中,其替代方案,纯文化传递,内在地都是一种稀罕且不稳定的状况。我们进一步展示出,有遗传基础的后成规则,预计存在于基因—文化协同进化之中,确实在人类身上发生了,并且对其经验分析必不可少的所有条件也都存在。
我们现在来到基因—文化协同进化理论的中心经验问题:在人类种群中,由遗传进化与文化进化的交互作用所产生的后成规则的严格性与特异性的程度。一个重要的考虑是,在基于后成约束文化根的两种表型的选择率方面,只在1%或更小的级别上有一点细微的差别,也可能造成遍及作为一个整体的种群的遗传进化,足以使个体发育偏向于有利的文化根。进一步说,遗传漂变能够在短至10代的时间里变得可检测。这一推论是以经典种群遗传学的二对等位基因模型与多基因模型为基础的(Crow and Kimura, 1970; Roughgarden,1979),而我们随后将直接以基因—文化协同进化的模型来证实它。它也被大量地记录在了关于定向(orientation)与其他初等形式的动物行为的选择实验中(Dobzhansky et al. ,1972)。其对于基因—文化协同进化的特殊含义将在第6章中得到探索。
相反地,在文化根的选择上,仅仅是一个很小的偏向,也足以产生出惊人的新型社会组织来。在《微观动机与宏观行为》( Micromotives and Macrobehavior )(1978)中,谢林(Schelling)提供了一些令人印象深刻的例子,关于以相对小的个人决定为基础的模式之上的大影响,包括个别司机减速10秒钟引起的交通堵塞、由无非是加入一个多数派的温和愿望而已导致的居住隔离、缘于成员利益小幅降低到一定阈值水平以下的机构倒闭,以此类推。看起来同样可能的是,在一个更大的可用集合(例如偏爱某些食品、服饰或婚姻安排的趋势)之外采用一个文化根子集的微弱的先天倾向,在某些条件下也有可能被放大成为文化进化中大得多的事件。例如,年轻男孩女孩早期性情上相对微小的先天差异(Blurton Jones and Konner,1973;Maccoby and Jacklin,1974),被扩大为所有社会中都一致的角色差异,包括少数中的极端男性支配(Rohner,1975;Draper,1976)。
这种放大的一个重要后果,是模式变动可由不同原因发起并沿独立路径实现的轻松性。由此,微观动机相对难以单独从一种宏观行为的知识中推断出来。快速的历史变迁经常被引用来作为人类社会行为中遗传约束缺席的证据(Allen et al. ,1976;M. Harris,1979)。然而,有所相关的并非文化进化的数量,而是其方向的一致性。模式对来自社会生物学理论细致预测的遵从,以及尤其是个体发育中变化与后成规则之间的因果关系,也是同样重要的。当来自发育心理学的精确信息被并入到基因—文化协同进化的模型之中时,社会科学意义重大的理论问题将变得更易于处理。
因此,文化研究应从关注后成规则调节遗传进化与文化进化之间关系的一类工作中受益。我们认为,这些规则的进化势必形成两种互补往复式的选择模式。第一种是 遗传同化 (genetic assimilation),其将构成如下事件序列:由基因型与正常物种环境相互作用产生的表型频率分布,为环境中的一个变化所扩展,创造出一种新颖的表型;于新环境中产生该表型的趋势,在先在的基因型之间变异;该新颖表型在新环境中占有遗传选择优势,导致一种使发育倾向于它的基因型的频率提高;在足够数量的几代以后,取决于选择的强度与传承的模式,作为一个整体的种群变得更容易发育该表型,而当该物种回到最初的环境时,该特征甚至会作为反应规范的一部分继续存在。尽管遗传同化迄今只在解剖学与生理学特征方面有所记录(Waddington,1962;Futuyma,1979;Milkman,1979),它应在行为表型,包括自身部分地构成了变动着的环境的文化创新方面同样可得。文化能力的遗传同化,预计可经由后成规则中的一个变化而发生,使得新颖文化根的传递更加可能(见图1.7)。
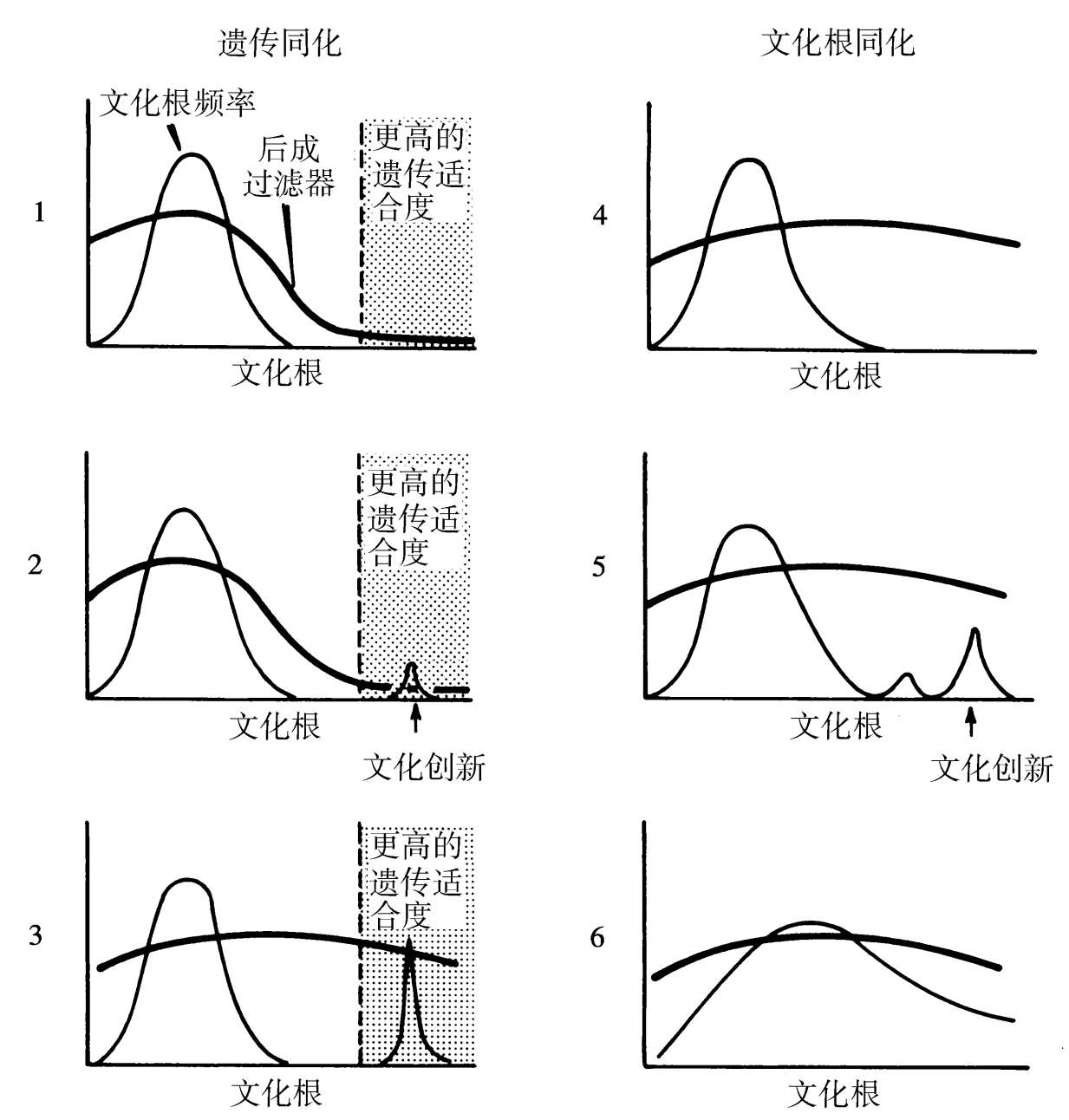
图1.7 基因—文化协同进化期间假设的遗传传递与文化传递互惠效应的简化表示。细线代表社会中可替代文化根的频率,而粗线表示学习、使用与传授特定文化根的倾向。 遗传同化 :一个文化根被发明出来,传达更大的遗传适合度(1,2);随后的自然选择改变着后成曲线而使传递更有可能发生(3)。 文化根同化 :后成规则已经普遍宽松,结果是文化根更容易被发明与传播(4—6)。尽管这两种同化模式在此特殊的图表中被顺次展示,它们也可能同时发生。进一步说,同化的方向能够被反转,结果将是后成规则的收窄与文化的贫瘠。
相反的过程,可称为 文化根同化 (culturgen assimilation):如果发育的灵活性足够大,某些新颖的文化根就有可能被发明与传播。换句话说,文化倾向于填补遗传决定后成规则所允许的空间。除此之外,文化在规则最偏爱它的地方的那些行为类别中,预计将是最为丰富的。我们可以期望文化堆积起来,成为围绕最受偏向性后成规则影响的习俗的节点,诸如乱伦回避、求偶以及群里群外之间的区别对待。最仪式化的文化形式,不会像许多社会科学家所思考的那样倾向于取代后成规则,而是会增强它们。
后成规则可能凭借处于少数有选择优势的新颖文化根的影响之下的遗传同化而进化,一种仍然接纳其他文化根的变化。换句话说,文化根同化跟在遗传同化之后。另一种可能,文化根同化也会跟在文化的贫瘠之后,而牵涉不到后成规则中的遗传变化。
如果用一种单程生命循环的抽象图来表示它们,基因—文化协同进化的初等特性能够被更清楚地理解到(见图1.8)。在每一个世代的进程之中,一个社会的文化根都要经过构成后成规则的“过滤器”来被清扫。如果该文化是由容易教与学的要素构成,并且并不是创新性的,它就将从一代到下一代地保持稳定。这些文化根对于该社会的成员们来说“感觉对”,经过许多世代,它们至多产生出一种相对弱强度的稳态化遗传自然选择,转而保持后成规则为近似恒定。如果另一方面,新文化根是通过创新或扩散引进的,它们将会被相对轻松地并入进来,并引起文化的扩张或转换,直到它们为后成规则所反对。这就是我们称之为文化根同化的过程。为后成规则所不喜的文化根将会被更快地清除,而开始遍及许多社会地只以更低的频率发生。这一过程预计会无限期地继续下去,除非新颖的文化根带来了更高的遗传适合度,在此情况下,后成规则自身可为在一个对于新颖文化根来说更为宽松的方向上经过数代时间的遗传进化所改变。这一形式的遗传同化在图中表现为后成过滤器的侵蚀。但相反的过程也可能发生:如果新文化根呈现出那种一致较低的适合度,后成规则将会被收紧。在比喻的图像中,过滤器现在向内生长。
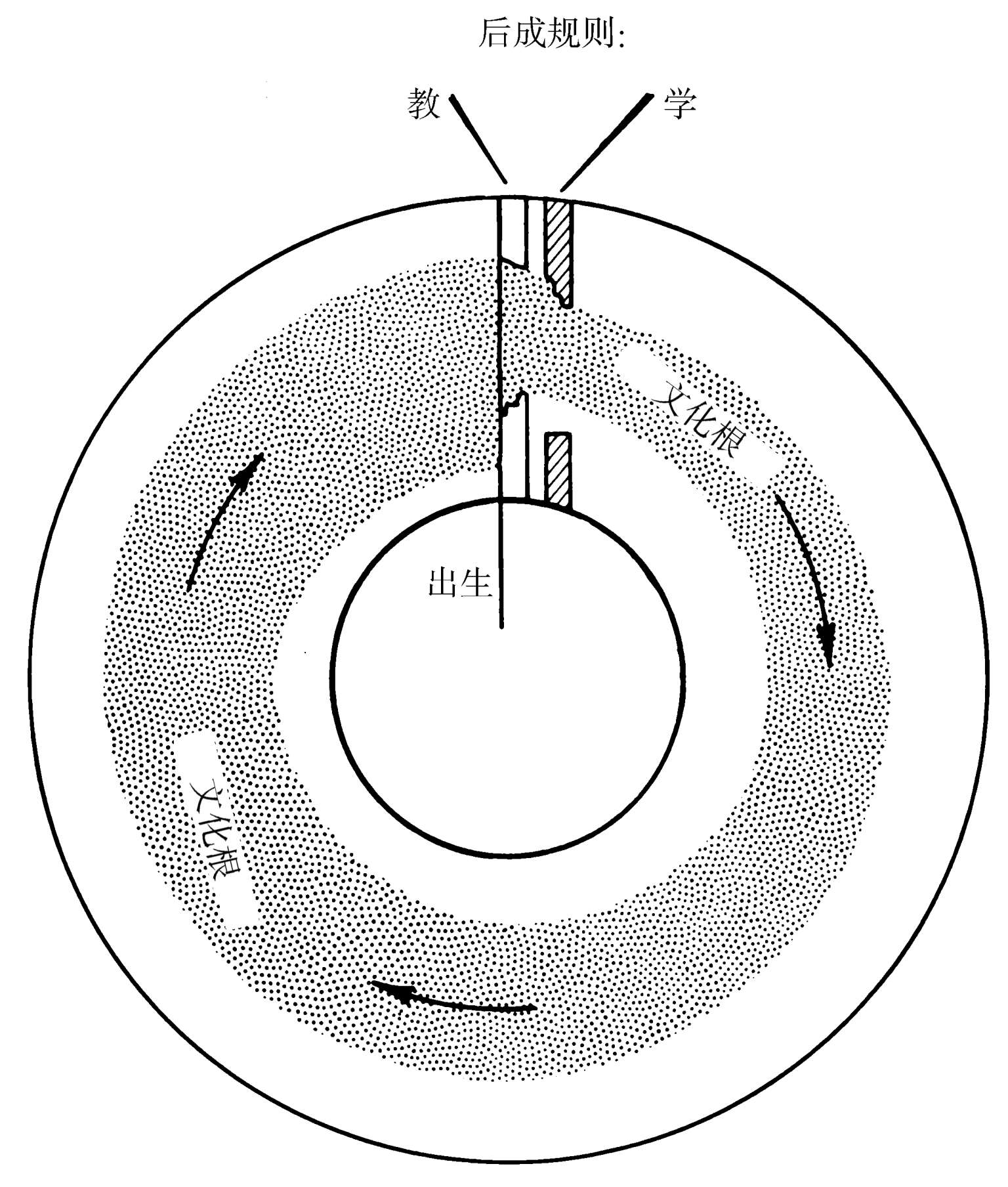
图1.8 基因—文化协同进化的抽象表示。圆环面包含一个社会成员的生命周期。文化根同化发生于文化根在个体的生命跨度之内被引入到社会之时。在此特例中,展示出一个非常大的数量。后成规则既影响个体传授特殊文化根的倾向,也影响学习它们的其他人,在图表中显示为一些过滤器。这些规则,影响着文化进化的速率与范围,放在这里作为童年早期的一次发生,但它们是能够多次且终生发生的。遗传同化发生于过滤器经过许多世代而被作用于后成规则之上的自然选择所改变之时。当新颖的文化根一致带来较高的遗传适合度,过滤器被侵蚀。当文化根降低遗传适合度,过滤器向内生长。
我们现在可以以一种更精确的形式重申这本书的目标:它是要用公式阐明一个理论框架,其将生成一个完全的模型光谱,从全遗传到全文化传递,涉及要么是整个物种仓库,要么是仓库内特殊的行为类别。这项计划建基于我们的假定之上,即人类文化传递终归是基因—文化传递,而人类文化进化只代表远大得多的可能传递史阵列的一个轨迹小集合。这传递空间,可通过一种比较社会理论的发展来让人理解,其中范围广泛的得到推论的非人类解也会被添加到传统的社会科学之中。
基因—文化协同进化理论也将大大扩展社会生物学的视野。动物学研究目前聚焦在了一个相对有限的现象集合之上,诸如亲缘选择、领地性与等级体系,这些最有可能将合作纳入动物的本能仓库之中。社会生物学还没有处理学习与认知或那些深度的、贯穿特征化人类行为发育的社会化的成果。基因—文化协同进化理论被设计来对付这些问题,并允许整个进化论进入到产生人类心灵及行为的机制的研究之中。正如我们将在即将到来的章节中展现的那样,它从生物学的基本规则中衍生出文化多样性的模式。并且因为人类社会化与认知实际上影响着每一种缺少反射性与自主性的行为特征,基因—文化论也扩展社会生物学远超出以动物为基础的研究的传统论题之外。它指示出人类后成规则大领域的存在,其大部分依然未被研究,它在文化进化引导方面的成果意义深远而仍然大大地不被赏识。