
下载掌阅APP,畅读海量书库
立即打开



任何物种的演化路径都是独特而无法预测的,无论是在其演化伊始还是演化轨迹行将到头之际。自然选择完全有可能把一个物种带到革命性的重大变化发生之前,却又改弦易辙。不过,我们至少可以判断有些演化轨迹在这个星球上有没有可能发生。比如昆虫能够变得非常微小,却绝无可能像大象一般大。猪也许可以变成水生动物,但它们的后代肯定不会飞。
我们可以用走迷宫的方式来说明一个物种可能经历的演化路径(见图3-1)。在实现一个重大进展的过程中,比如真社会性的产生,每一次遗传变化就相当于迷宫中的一个转角,要么使得达成这一进展的可能性更低甚至使之成为不可能,要么离这一进展更近一步。最开始的几步,也就是仍然存在一些其他选项的情况下,要走通迷宫还有很长的路,离终点也远得很,完成的可能性非常低。到最后几步,距离终点已经不远,成功的可能性就变得大多了。在这一过程中,迷宫本身也受到演化的控制。旧通道(生态位)会关闭,新通道会打开。迷宫结构在某种程度上依赖于迷宫里面穿行的每一个物种。
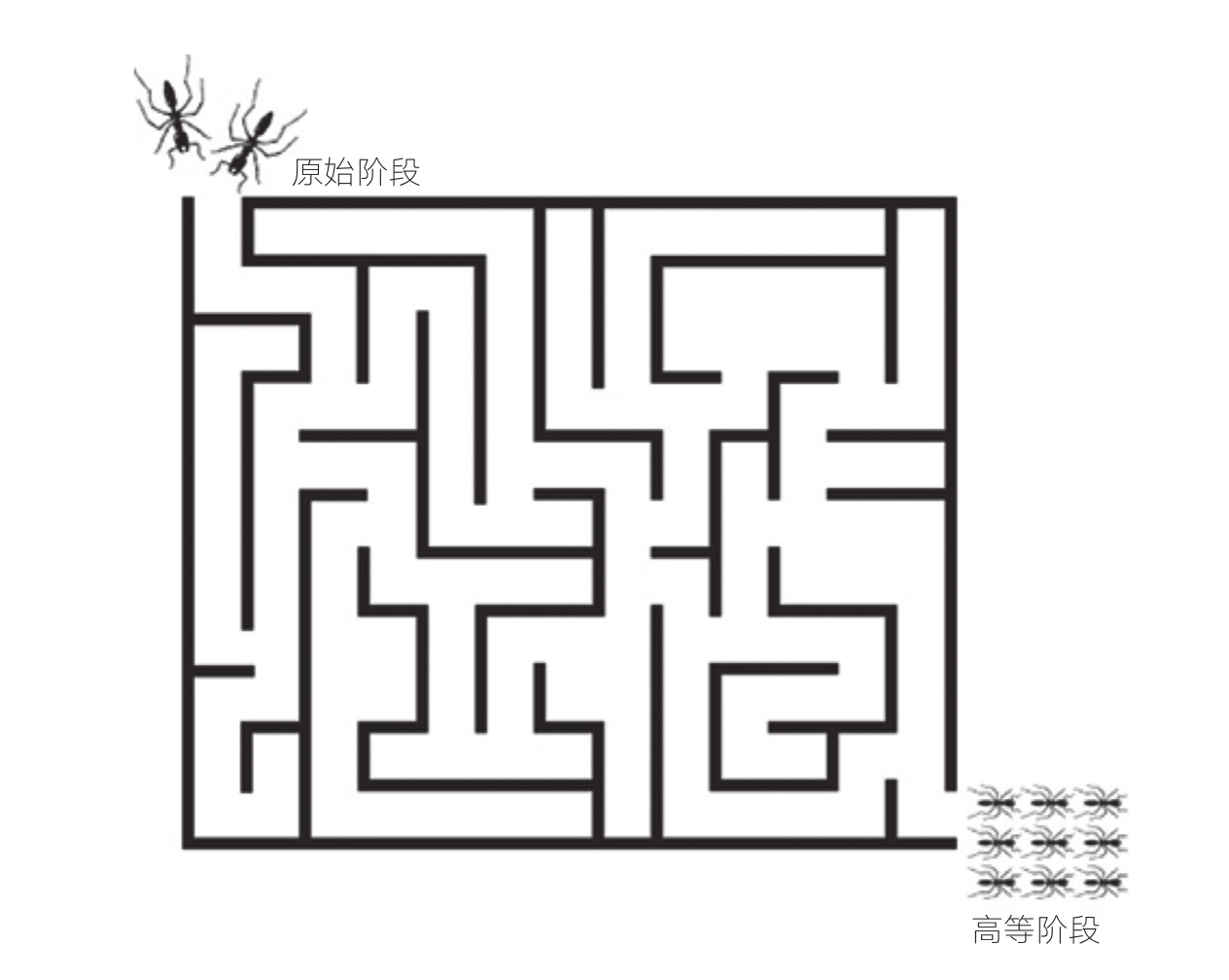
图3-1 演化迷宫
一个物种的演化图景可以看作一个由环境呈现的迷宫,伴随着迷宫自身的演变,各种机会不断消失又出现。图3-1表示的是从原始社会性到高等社会性的演化路径。
每一场演化都是一场概率意义上的赌博,涉及一代又一代的个体,必然会有大量个体生生死死。不过个体数量并非大得不可估量,至少我们可以大致估算一下数量级。从1亿年前我们的原始哺乳动物祖先开始算起,到第一支智人种的出现,整个过程所需要的个体总数大约为1 000亿。这些祖先就这么生来又死去,才有了今天的我们。
演化迷宫里还有许多其他物种在不断经历着衰落和消亡,其中大部分平均每一代都有几千个具备繁殖能力的个体。在人类的一长串祖先中,只要任何一支走入死胡同,人类史诗般的演化就会戛然而止。我们的前人类祖先既非上天指派,也不是格外伟大,它们只不过极其幸运。
近年有一些研究综合了多个不同的科学学科,正在合力阐明人类起源的几个阶段,为困扰科学界和哲学界的“人类独特性”问题多少提供了一些解释。如果回过头从人类起源开始审视,一直到人类取得现有的地位,其间每个阶段都可以被理解为演化的预适应。我这么说并不是指人类的各个祖先物种是有了某种引导才有现在的结局。相反,人类演化的每一步就其自身而言都是一种适应,是自然选择对物种当时当地所处环境的反应。
第一项预适应是前面提到的大体形以及行动范围的相对固定,这些特性预先决定了哺乳动物不同于社会性昆虫的演化轨迹。人类演化进程表中的第二项预适应发生在距今8 000万~7 000万年前,早期灵长目动物适应了树上生活。这种变化演变出的最重要特征是适合抓握的手脚,而且手脚的形状和肌肉不单适应于抓握树枝帮助支撑自身体重,还更有利于身体在树枝间摇荡。与此同时,他们还演化出了可与其他手指对握的拇指和大脚趾,以提高抓握的效果。进一步增强抓握能力的演化特征是变平的指甲和趾甲,这和其他大部分树栖哺乳动物那种尖利下弯的爪子相反。此外,人类的手掌和脚掌覆有掌纹,也能辅助抓握;掌上的压力感受器能够增强触觉感受。因此,带有这些“装备”的早期灵长目动物能用手采摘、扒拉水果块并把籽给弄出来,用指甲切割、刮擦手里抓着的东西。它们手里抓着食物,后腿用来移动,能带着食物跑相当长一段距离,而不必像猫狗那样动用上下颌咬住食物,也不必像筑巢的鸟那样靠反哺给小鸟喂食。
前人类灵长目具有相对复杂的生活方式和灵活的进食行为,栖息地植被开阔,兼具一定的广度和高度(见图3-2)。也许是作为对上述条件的适应,前人类灵长目演化出了更大的大脑。同理,相比大多数其他哺乳动物,它们变得更为依赖视觉,而较少依赖嗅觉。它们获得了具备色感的大眼睛,其位于头的前部,产生的双目视觉可以形成更好的深度感。前人类灵长目行走时两腿不会走成两条平行线,而是左右交替几乎成一直线,一只脚在另一只脚的正前方。并且,它们每一胎繁殖的后代数量较少,后代发育需要的时间较长。

图3-2 一只黑猩猩两足行走在塞内加尔方戈里的稀树草原中
图片来源:Mary Roach, “Almost Human,” National Geographic , April 2008, p. 128. Photograph by Frans Lanting. Frans Lanting / National Geographic Stock.
当这群奇特的树栖生物中有一支在非洲发展至适应陆地生活时,又一项预适应产生了——演化迷宫中的又一个幸运转角:采用两只后足行走,用双手来完成其他任务。现存的两个猩猩种类,黑猩猩和倭黑猩猩,也是在系统发育上与人类亲缘关系最近的物种,它们与人类在大约同一时间朝相同的方向行进了相当一段距离。如今在陆地上,它们也时常举起双臂用后腿行走和奔跑,甚至能够制作原始的工具(见图3-3)。

图3-3 一只黑猩猩坐在白蚁蚁丘上
这里是前人类的栖息地。在这里,它们也使用粗糙的工具。
图片来源:W. C. McGrew, “Savanna chimpanzees dig for food,” Proceedings of the National Academy of Sciences , U.S.A. 104[49]: 19167-19168 [2007]. Photograph by Paco Bertolani, Leverhulme Centre for Human Evolutionary Studies.
前人类在演化路上与黑猩猩分道扬镳后,就形成了现在被称为南方古猿的物种。它们朝两足行走的方向走得更远,整个身体也发生了相应的变化:腿变长变直了,细长的双足会使身体在移动时摇晃,由于内脏不再像猿猴似的垂挂在水平的躯干下面,而是压向两条腿,于是为了支撑内脏,骨盆变成了浅碗状。
两足行走这个关键变化极有可能是南方古猿全面获胜的原因,这一改变至少让它们有了多样化的体形、颚肌和牙列。大约200万年前,那段时间非洲大陆存在着至少三种南方古猿。它们的身体比例、直立姿态、颤巍巍竖在身体上面的脑袋以及用来跑跳的两条长腿,看起来都和现代人类相去甚远。几乎可以肯定它们是集合成一小群一小群迁徙的,就像游牧民族一样。它们的大脑并不比黑猩猩的大,却最终演化出了第一批智人。在物种演化的过程中,机会源自多样性,南方古猿发现了这一点。
南方古猿及其后代物种构成的人属都生活在有利于直立行走的环境里。它们从不像黑猩猩等现代猿类那样跖行,即采用两手蜷握充当前脚的行走方式,而是两臂悬在身体两侧。这种新的南方古猿式行走方法用最少的能量消耗换来了速度的提升,但同时也给背部和膝盖带来了问题。为了承托细脖子上的那颗沉重的脑袋(见图3-4),尽力保持平衡,它们也要面对更大的风险。

图3-4 地猿(重建图)
在埃塞俄比亚阿瓦什中部地区发现的地猿谱系化石距今440万年,是已知最古老的现代人类的两足祖先。它依靠细长的后腿行走,同时保留适合树上生活的长臂。
图片来源:Jamie Shreeve, “The evolutionary road,” National Geographic , July 2010, pp. 34-67. Painting by Jon Foster. Jon Foster / National Geographic Stock.
对于身体原为适应树上生活而打造的灵长类动物来说,双足行走让它们能够快速奔跑,可它们却跑不过想要捕猎的四足动物。羚羊、斑马、鸵鸟等动物都能轻轻松松地在短距离内超过它们。在狮子等食肉短跑健将数万年的追赶下,这些猎物都被训练成了百米冠军。不过,早期人类虽然在短跑比赛中跑不过这些动物健将,但至少还能在马拉松比赛中战胜它们。到一定程度后,人类成了长跑选手。人类只要跟在猎物后头跑啊跑,跑个几千米,跑得猎物精疲力尽时就能追上它们了。前人类每跑一步都要靠足底的跖骨球用力,还要保持节奏稳定,此外还演化出了很强的携氧能力来适应长跑。经过一段时间后,它们的毛发变少了,除了头部、耻骨和分泌激素的腋下,身体其他部位的毛发都脱落了。相较之下,汗腺则增加了,全身分布的汗腺让裸露的身体表面能够更快地散热。
伯恩德·海因里希(Bernd Heinrich)是一位杰出的生物学家,也是破过纪录的超长距离跑步名将,他在《与羚羊赛跑》(
Racing the Antelope
)一书中详尽地阐述了他的马拉松理论。他引用2000年获得25千米跑美国国家冠军肖恩·方德(Shawn Found)的话来说明长久奔跑带给人的原始愉悦感:“奔跑让你重温捕猎的过程。你追赶着那些比你冲刺得快的猎物,一追30英里
 ,最后追到并把猎物带回村庄,这就是跑步,一件无比美妙的事。”
,最后追到并把猎物带回村庄,这就是跑步,一件无比美妙的事。”
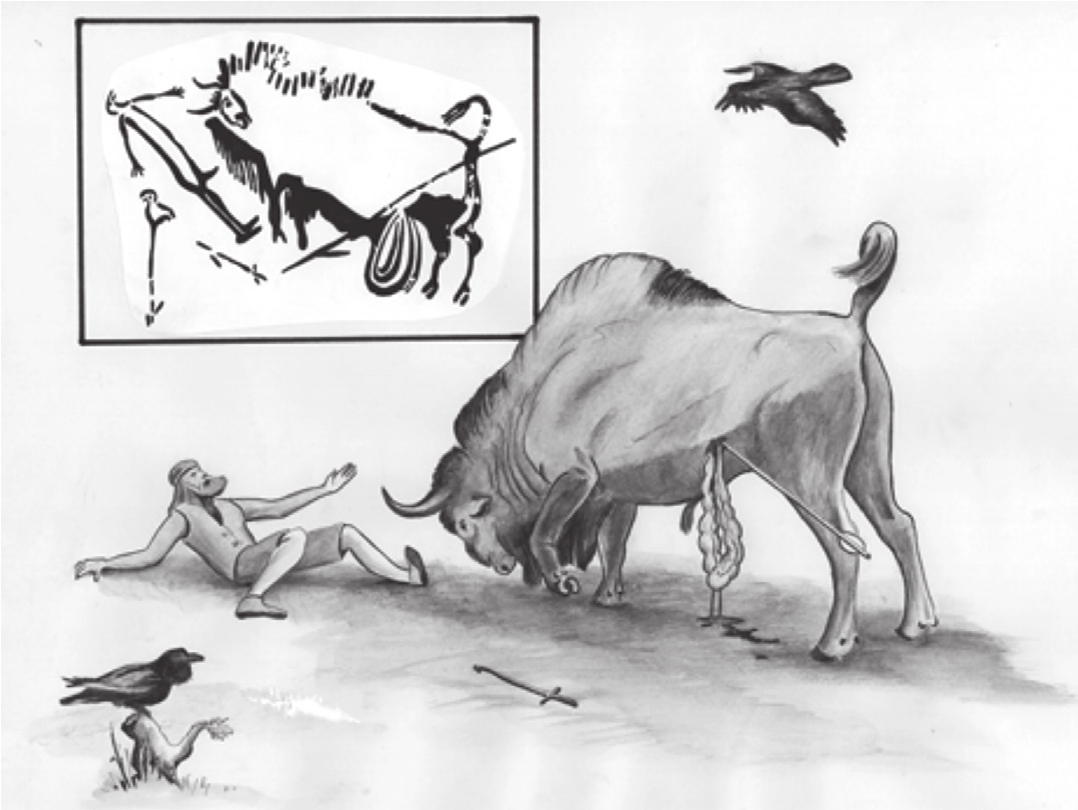
图3-5 拉斯科洞穴壁画中的捕猎场景
在史前时期,狩猎是一种具有高度适应性的、危险的行为。这幅插图是旧石器时代拉斯科洞穴壁画的一部分,描绘了一头中箭的野牛扑向一名倒下的猎人,而一只乌鸦(一种跟随猎人的常见食腐动物)在附近徘徊。图片来源:R. Dale Guthrie in The Nature of Paleolithic Art, Chicago: University of Chicago Press, 2005.
与此同时,前人类祖先的前肢发生了改变,可以灵活地操纵物体。手臂,尤其是雄性的手臂,变得擅长投掷,比如石块以及后来的矛等,到这个时候前人类才终于能够实施远距离猎杀。这种技能让它们在与装备较差的群体发生冲突时占据了优势。
现在的普通黑猩猩中至少有一个群体学会了投掷石块。这种行为或许是由某个个体偶然为之,看上去却像一场文化革新。但不可思议的是,没有任何黑猩猩的投掷能力可与现代人类中的运动员媲美。没有黑猩猩能以约每小时140千米的时速扔出石块或将长矛掷出一个足球场远的距离,年轻力壮的黑猩猩即便经过训练也掌握不了人类儿童就能掌握的投掷技巧。早期人类利用天生的肢体通过投掷的方法抓捕猎物、驱逐敌人,并取得了决定性的优势。在今天的考古现场,矛尖和箭头属于最早出现的人工制品。
前人类时期的环境对于产生两足行走的动物和它们的马拉松后代来说十分理想。在关键的演化阶段,非洲撒哈拉以南大部分地区非常干旱,雨林面积减少,雨林逐渐撤向赤道带,只剩在非洲北部的零星据点。交错的热带稀树草原和干旱的草原覆盖了非洲大陆总面积的很大一部分。前人类和智人在外面觅食时,可以站着并越过低矮的植被用目光搜寻猎物,防备捕食者(见图3-6)。金合欢等优势树种相对较矮,树冠部的枝杈向下延展到地面,很容易攀爬,这些特点都对二足动物十分有利。坦桑尼亚的塞伦盖蒂草原、肯尼亚的安博塞利、莫桑比克的戈龙戈萨以及东非其他一些国家公园现在仍然保留着类似的环境结构。相比于非洲撒哈拉以南的其他栖息地,诗人和游客都更爱这样的环境。不过我稍后会解释,他们可能是被一种本能感动了,而这种本能正是由他们生活在此的祖先经过数百万年演化而来的。

图3-6 在卡拉哈里南部的草原上觅食的布须曼人(也叫桑人或巴萨尔瓦人)
这一景象可能与6万年前在同一地区经常发生的景象没有太大区别。
图片来源:Stephan C. Schuster et al., “Complete Khoisan and Bantu genomes from southern Africa,” Nature 463: 857, 943-947 (2010). Photo © Stephan C. Schuster.
人类的发源地并非树冠高耸、腹地幽暗的茂密雨林,也不是相对平淡无奇的草原和沙漠。人类诞生于稀树草原,那儿交杂着各种不同的局部生境。
在通往真社会性的道路上,接下来的步骤是掌握火的用法。雷电引发地表火在今天的非洲草原和森林也是司空见惯的事。林间溪流环绕的林地和动辄洪水泛滥的潮湿洼地将火势削弱后,林地的下层灌丛就会变得茂密,长成易燃物。雷电和地表火蔓延会引发野火,这不仅会引燃四周的地表植被,还会向上烧到周围树林的树冠层。一些动物,尤其是动物中的老幼病残会在火海中丧生。四处游荡的前人类肯定会注意到野火是获得食物来源的重要途径。并且,它们还会发现有些动物尸体被烤熟后,肉变得容易撕咬,吃起来更方便了。
澳大利亚土著不仅到了现代仍在接受自然的这份馈赠,还会用树枝做成的火把传播火种。前人类祖先有没有可能也是这么做的呢?对于这些早期的情形,我们无从得知,但是在通往现代人类境况的曲折道路中,人属在历史早期掌握对火的使用无疑是一个重大事件。
而火永远不会被昆虫和其他陆生无脊椎动物所用。因为它们体形太小,无法点燃火种,也无法在不伤及自身的情况下携带燃烧物。当然水生动物也不会学习用火,不管它们的体形有多大、智力有多高以及其他天性如何。与人类拥有同等智力的物种只能在陆地上出现,无论是地球的陆地还是其他什么宜居星球的陆地。就算在幻想世界中,美人鱼和海神也得先在陆地上演化出相当的智能才会返回它们统治的水域。
接下来是人类真社会性起源的决定性一步,根据研究者从其他动物那里采集的证据,营地小规模的聚集开始出现。集体由数代同堂的大家庭组成,参照现代狩猎采集社会的人员结构,我们会发现其中也有通过异族婚姻交换而来的外族妇女。
大量考古学证据告诉我们,早期非洲智人和他们的姊妹种欧洲尼安德特人,以及他们的共同祖先直立人,都有营地。因此这一现象可以追溯到至少100万年前。我们认为建立营地是发展出真社会性的关键适应性改变,这有一个先验的理由,即营地本质上就是人类所建的巢穴。所有发展出真社会性的物种,无一例外会先筑巢来抵御敌人。它们和已经为人所知的祖先一样,在巢穴中哺育后代,离巢觅食并把多余的食物带回来和同伴分享。在原始白蚁、粉蠹虫、棉蚜和蓟马中,虽然这种行为发生了一些变化——巢穴本身就是它们的食物,但基础配置没有变,仍遵从着巢穴在真社会性演化中占据重要位置这一生物学原则。
晚成鸟,即雏鸟无独立生活能力需要亲鸟喂养的鸟类,也有类似的预适应。少数几种鸟中,刚成年的小鸟会在巢中停留一段时间,帮助亲鸟一起照顾弟弟妹妹。但没有任何鸟类继续演化出完善的真社会性。它们有的只是喙和爪子,从未具备使用更复杂的工具的能力,也完全不曾掌握火的用法。狼和非洲野犬像黑猩猩、倭黑猩猩一样合作狩猎(见图3-7)。非洲野犬也会挖洞,可供一到两头雌性伴侣生下一大窝幼崽。犬类中有的成员会出去狩猎并把一部分猎物带回巢穴给母狗和小狗吃,其他成员则留守看家。这种引人瞩目的犬类,尽管获得了罕见且极其不容易的预适应,也仍未能达到真正的真社会性、具备专职工犬或猿类的智力。它们没有能力制造工具,缺少适合抓握的爪与末端柔软的指头。它们依然四脚着地,依赖于裂齿和覆毛的爪子。

图3-7 非洲野犬
图片来源:E. O. Wilson, Sociobiology (Cambridge, MA: Harvard University Press, 1975), pp. 510-511. Drawing by Sarah Landry.