
下载掌阅APP,畅读海量书库
立即打开



耳分为外耳(external ear)、中耳(middle ear)和内耳(inner ear)三部分。颞骨内包含了外耳道的骨部、中耳、内耳和内耳道(图2-1-1)。
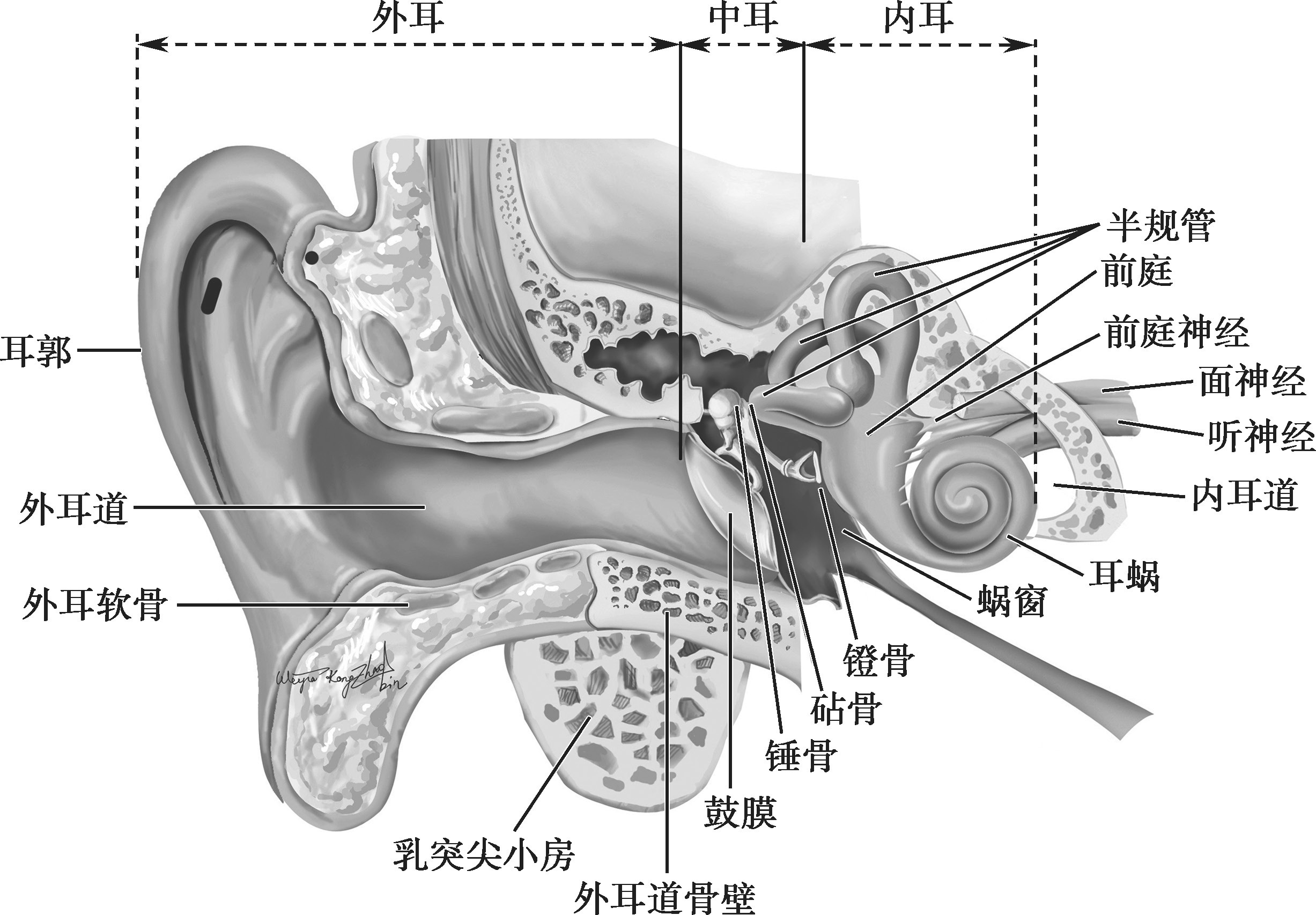
图2-1-1 外耳、中耳、内耳关系示意图
颞骨(temporal bone)是解剖结构最为复杂的人体结构之一,位于颅骨两侧,镶嵌在顶骨、蝶骨、颧骨和枕骨之间,参与构成颅中窝和颅后窝的侧壁和底壁。颞骨为复合骨,以外耳道为中心可将颞骨分为5部分,即鳞部、鼓部、乳突部、岩部和茎突。
外耳(external ear)包括耳郭和外耳道。
耳郭(auricle)内含弹性软骨支架,外覆皮肤。一般与头颅约成30°角,左右对称,分前(外)面和后(内)面。耳郭前(外)面凹凸不平,主要表面标志有:耳轮、耳轮脚、耳轮结节、三角窝、舟状窝或耳舟、耳甲艇、耳甲腔、耳屏、对耳屏和耳屏间切迹等。
外耳道(external auditory meatus)起自耳甲腔底,向内止于鼓膜,由软骨部和骨部组成,略呈S形弯曲,长2.5~3.5cm。成人外耳道外1/3为软骨部,内2/3为骨部。新生儿外耳道软骨部与骨部尚未完全发育,由纤维组织所组成,故外耳道较狭窄而软组织塌陷。1岁以下的婴儿外耳道几乎为软骨所组成。外耳道有两处较狭窄,一为骨部与软骨部交界处,另一为骨部距鼓膜约0.5cm处,后者称为外耳道峡(isthmus)。发育正常的成人外耳道外段向内、向前微向上,中段向内、向后,内段向内、向前而微向下;故在检查成人外耳道深部或鼓膜时,需将耳郭向后上提起,使外耳道成一直线。
中耳(middle ear)介于外耳和内耳之间,是位于颞骨中的不规则含气腔和通道,包括鼓室、咽鼓管、鼓窦及乳突4部分。
鼓室(tympanic cavity)为颞骨内最大的不规则含气腔,位于鼓膜与内耳外侧壁之间。鼓室前方经咽鼓管与鼻咽相通,后方经鼓窦入口与鼓窦及乳突气房相通。以鼓膜紧张部的上、下缘为界,将鼓室分为3部:①上鼓室(epitympanum),或称鼓室上隐窝(attic),位于鼓膜紧张部上缘平面以上的鼓室腔;②中鼓室(mesotympanum),位于鼓膜紧张部上、下缘平面之间,即鼓膜紧张部与鼓室内壁之间的鼓室腔;③下鼓室(hypotympanum),位于鼓膜紧张部下缘平面以下,下达鼓室底。鼓室的上下径约15mm,前后径约13mm;内外径在上鼓室约6mm,下鼓室约4mm,中鼓室于鼓膜脐与鼓岬之间的距离最短,仅约2mm。鼓室的容积为1~2mL。鼓室内有听骨链、肌肉及韧带等。鼓室腔内为黏膜所覆盖,覆于鼓膜内侧面、鼓岬后部、听骨链、上鼓室、鼓窦及乳突气房者为无纤毛扁平上皮或立方上皮,余为纤毛柱状上皮。
鼓室内容物包括听小骨、韧带和肌肉。
为人体中最小的一组骨骼,包括锤骨、砧骨和镫骨。三者相互以关节连接形成链状,称为听骨链(ossicular chain)。听骨链位于鼓膜和前庭窗之间,通过其杠杆作用,将鼓膜感受到的振动传入内耳。
1)锤骨(malleus):
形如锤,位于鼓室中部和最外侧,长约8~9mm,有头、颈、外侧突(短侧突)、前突(长突)和柄。锤骨柄位于鼓膜黏膜层与纤维层之间,锤骨头位于上鼓室,其头的后内方有凹面,与砧骨体形成砧锤关节。
2)砧骨(incus):
分为体、长脚和短脚,长脚约7mm长,短脚约5mm长。砧骨体位于上鼓室后方,其前与锤骨头相接形成砧锤关节。短脚位于鼓窦入口底部,其尖端借韧带附于砧骨窝内。长脚位于锤骨柄之后、与锤骨柄平行,末端内侧有一膨大向内的突起名豆状突(lenticular process),后者有时与长脚末端不完全融合,故又名第四听骨。豆状突与镫骨头形成砧镫关节。
3)镫骨(stapes):
形如马鞍,分为头、颈、前脚、后脚和足板(foot plate),高3~4mm。镫骨头与砧骨长脚豆状突相接。颈甚短,其后有镫骨肌腱附着。前脚较后脚细而直,两脚内面各有深沟。两脚与镫骨足板之间的空间称为闭孔。镫骨足板呈椭圆形,长3mm,宽1.4mm,借环状韧带(annular ligament)连接于前庭窗。
有锤骨上韧带、锤骨前韧带、锤骨外侧韧带、砧骨上韧带、砧骨后韧带和镫骨环状韧带,将听骨固定于鼓室内。
1)鼓膜张肌(tensor tympani muscle):
起自咽鼓管软骨部、蝶骨大翼和鼓膜张肌管壁等处,其肌腱向后绕过匙突呈直角向外止于锤骨颈下方,由三叉神经下颌支的一小支司其运动;此肌收缩时牵拉锤骨柄向内,增加鼓膜张力。
2)镫骨肌(stapedius muscle):
起自鼓室后壁锥隆起内,其肌腱自锥隆起穿出后,向前下止于镫骨颈后方,由面神经的小支司其运动;此肌收缩时可牵拉镫骨头向后,使镫骨足板以后缘为支点,前缘向外跷起,以减少内耳压力。
动脉血液主要来自颈外动脉。上颌动脉的鼓室前动脉供应鼓室前部,耳后动脉的茎乳动脉供应鼓室后部及乳突,脑膜中动脉的鼓室上动脉及岩浅动脉供应鼓室盖及内侧壁,咽升动脉的鼓室下动脉供应鼓室下部及鼓室肌肉;颈内动脉的鼓室支供应鼓室前壁。鼓膜外层由上颌动脉耳深支供给,鼓膜内层由上颌动脉鼓前支和茎乳动脉分支供给。鼓膜的血管主要分布在松弛部、锤骨柄和紧张部的周围。故当鼓膜发炎时,充血自鼓膜松弛部开始,继则延伸至锤骨柄及鼓膜其他部分。静脉流入翼静脉丛和岩上窦。
主要为鼓室丛与鼓索。
1)鼓室丛(tympanic plexus):
由舌咽神经的鼓室支及颈内动脉交感神经丛的上、下颈鼓支组成,位于鼓岬表面,司鼓室、咽鼓管及乳突气房黏膜的感觉。
2)鼓索(chorda tympanic nerve):
自面神经垂直段的中部分出,在鼓索小管内向上向前,约于锥隆起的外侧进入鼓室,经砧骨长脚外侧和锤骨柄上部内侧,相当于鼓膜张肌附着处下方,向前下方经岩鼓裂出鼓室,与舌神经联合终于舌前2/3处,司味觉。
咽鼓管(eustachian tube)位于颞骨鼓部与岩部交界处,颈内动脉管的外侧,上方仅有薄骨板与鼓膜张肌相隔,为沟通鼓室与鼻咽的管道。成人咽鼓管全长约35mm,外1/3为骨部,内2/3为软骨部。咽鼓管鼓室口位于鼓室前壁上部,咽鼓管咽口位于鼻咽侧壁,下鼻甲后端的后上方。自鼓室口向内、向前、向下达咽口,故咽鼓管与水平面约成40°角,与矢状面约成45°角。骨部为开放性管腔,内径最宽处为鼓室口,越向内越窄。骨与软骨部交界处最窄,称为峡,长约2mm,内径约1mm。自峡向咽口又逐渐增宽。软骨部的后内及顶壁由软骨板构成,前外壁系由黏膜和肌膜组成,静止状态时软骨部闭合成一裂隙。由于腭帆张肌、腭帆提肌、咽鼓管咽肌起于软骨壁或结缔组织膜部,前两肌止于软腭,后者止于咽后壁,故当张口、吞咽、哈欠、唱歌时借助上述三肌的收缩,可使咽口开放,以调节鼓室气压,保持鼓膜内、外压力平衡。咽鼓管黏膜为假复层纤毛柱状上皮,纤毛运动方向朝向鼻咽部,可使鼓室分泌物得以排出;又因软骨部黏膜呈皱襞样,具有活瓣作用,故能防止咽部液体进入鼓室。成人咽鼓管咽口约高于咽口2~2.5cm;儿童咽鼓管接近水平,管腔较短、近成人的一半,且内径较宽,故儿童的咽部感染较易经此管侵入鼓室。
鼓窦(tympanic antrum)为鼓室后上方的含气腔,内覆有纤毛黏膜上皮,前与上鼓室、后与乳突气房相连,出生时即存在。鼓窦向前经鼓窦入口与上鼓室相通,向后下通乳突气房;上方以鼓窦盖与颅中窝相隔,内壁前部有外半规管隆突,后壁借乳突气房及乙状窦骨板与颅后窝相隔,外壁为乳突皮质,相当于外耳道上三角区(Macewen三角)。成人鼓窦的大小、形状因人而异,并与乳突气化的程度有直接关系。
乳突(mastoid process)为鼓室和鼓窦的外扩部分。乳突气房分布范围因人而异,发育良好者,向上达颞鳞,向前经外耳道上部至颧突根内,向内伸达岩尖,向后伸至乙状窦后方,向下可伸入茎突。根据气房发育程度,乳突可分为4种类型:①气化型,乳突全部气化,气房较大而间隔的骨壁较薄,此型约占80%;②板障型,乳突气化不良,气房小而多,形如头颅骨的板障;③硬化型,乳突未气化,骨质致密,多由于婴儿时期鼓室受羊水刺激、细菌感染或局部营养不良所致;④混合型,上述3型中任何2型同时存在或3型俱存者。乳突在初生时尚未发育,呈海绵状骨质,随着年龄增长逐渐发育,6岁左右儿童的气房已有广泛的延伸,最后形成许多大小不等、形状不一、相互连通的气房,内有无纤毛的黏膜上皮覆盖。
内耳(inner ear)又称迷路(labyrinth),埋藏于颞骨岩部,结构复杂而精细,内含听觉和前庭器官。
耳蜗(cochlea)位于前庭的前部,形似蜗牛壳,主要由中央的蜗轴和周围的骨蜗管组成。骨蜗管(蜗螺旋管)旋绕蜗轴2.5~2.75周。蜗底向后内方,构成内耳道底。蜗顶向前外方,靠近咽鼓管鼓室口。蜗底至蜗顶高约5mm,蜗底最宽直径约9mm,蜗轴呈圆锥形。骨蜗管内共有3个管腔:上方名前庭阶,自前庭窗开始;中间为膜蜗管,又名中阶,系膜迷路;下方名鼓阶,起自蜗窗,为蜗窗膜(第二鼓膜)所封闭。骨螺旋板顶端形成螺旋板钩,蜗轴顶端形成蜗轴板;螺旋板钩、蜗轴板和膜蜗管顶盲端共围成蜗孔。前庭阶和鼓阶的外淋巴经蜗孔相通。蜗神经纤维通过蜗轴和骨螺旋板相接处的许多小孔到达螺旋神经节。耳蜗底转的最下部、蜗窗附近有蜗水管开口,其外口在岩部下面颈静脉窝和颈内动脉管之间的三角凹内,鼓阶的外淋巴经蜗水管与蛛网膜下腔相通。
膜蜗管(membranous cochlear duct)位于骨螺旋板与骨蜗管外壁之间,为耳蜗内螺旋形的膜质管道,在前庭阶与鼓阶之间,又名中阶,内含内淋巴。此乃螺旋形的膜性盲管,两端均为盲端。膜蜗管的横切面呈三角形,有上、下、外三个壁:上壁为前庭膜,起自骨螺旋板,向外上止于骨蜗管的外侧壁;外壁为螺旋韧带,上覆假复层上皮,内含丰富的血管,名血管纹;下壁由骨螺旋板上面的骨膜增厚形成的螺旋缘和基底膜组成。基底膜起自骨螺旋板的游离缘,向外止于骨蜗管外壁的基底膜嵴。位于基底膜上的螺旋器又名Corti器,是听觉感受器的主要部分。基底膜在蜗顶较蜗底宽,即基底膜的宽度由蜗底向蜗顶逐渐增宽,而骨螺旋板及其相对的基底膜嵴则逐渐变窄。
在螺旋器中的螺旋隧道(Corti tunnel)、Nuel间隙及外隧道的间隙中,充满着和外淋巴性质相仿的液体,称Corti淋巴。此系通过骨螺旋板下层中的小孔及蜗神经纤维穿过的细孔与鼓阶的外淋巴相交通。膜迷路的其他间隙均充满内淋巴。因此,螺旋器听毛细胞的营养来自Corti淋巴(其离子成分与外淋巴相似),而囊斑及壶腹嵴感觉细胞的营养均来自内淋巴。螺旋器(Corti器):位于基底膜上,自蜗底至蜗顶全长约32mm,由内、外毛细胞,支持细胞和盖膜等组成。靠蜗轴侧有单排内毛细胞,其外侧有3排或更多的外毛细胞,这些是听觉感受细胞。内毛细胞呈烧瓶状,约有3 500个,外毛细胞呈试管状,约有12 000个。
面神经(facial nerve)是人体中居于骨管中最长的脑神经,也是容易发生麻痹的神经之一,含有运动纤维、感觉纤维及副交感纤维,以运动纤维为主。因此,面神经从中枢到末梢之间的任何部位受损,皆可导致部分性或完全性面瘫。面神经的运动神经核位于脑桥下部,此核向上通往额叶中央前回下端的面神经皮层中枢。部分面神经核接受来自对侧大脑运动皮层的椎体束纤维,从这部分面神经核发出的运动纤维支配同侧颜面下部的肌肉。其余部分的面神经核接受来自两侧大脑皮质的锥体束纤维,从此发出的运动纤维支配额肌、眼轮匝肌、皱眉肌。因此,当一侧脑桥以上到大脑皮质之间受损时,仅引起对侧颜面下部肌肉瘫痪,而皱眉及闭眼功能均存在。面神经运动核与三叉神经、视神经及蜗神经核有联系,因此能使某些肌肉完成一定的反射性收缩,如机体受到触觉、视觉或听觉刺激时发生眨眼反射;一定强度的声刺激可引起两侧镫骨肌的反射性收缩。面神经的运动纤维绕过展神经核后,在脑桥下缘穿出脑干。
上起额叶中央前回下端的面神经皮层中枢,下达脑桥下部的面神经运动核。
面神经根在脑桥中离开面神经核后,绕过展神经核至脑桥下缘穿出。
面神经离开脑桥后,跨过脑桥小脑三角,会同听神经抵达内耳门。此段长13~14mm,虽不长,但可被迫扩展到5cm而不发生面瘫。
从内耳门至内耳道底部的一段,长约10mm。此段面神经由内耳门进入内耳道,同听神经到达内耳道底。
面神经由内耳道低的前上方进入面神经管,向外于前庭与耳蜗之间到达膝神经节(geniculate ganglion)。此段最短,长2.25~3mm。
鼓室段又名水平段,自膝神经节起向后并微向下,经鼓室内壁的骨管,在前庭窗下方、外半规管下方,到达鼓室后壁锥隆起平面。此段长约11mm。此处骨壁最薄,易遭病变侵蚀或手术损伤,也可先天发育缺失;亦可将此段分为鼓室段(自膝神经节到外半规管下方)与锥体段(自外半规管下方到锥隆起平面)。
乳突段又称垂直段。自鼓室后壁锥隆起高度下达茎乳孔,长约16mm。此段部位较深,在成人距乳突表面大多超过2cm。颞骨内面神经全长约为30mm;其中自膝神经节到锥隆起长约11mm,自锥隆起到茎乳孔长约16mm。
面神经出茎乳孔后,在茎突的外侧向外、向后行进入腮腺。主干在腮腺内分为上支与下支,两者弧形绕过腮腺岬部后又分为5支;各分支间的纤维互相吻合,最后分布于面部表情肌群。
自膝神经节的前方分出,经翼管神经到蝶腭神经节,分布到泪腺及鼻腔腺体。
自锥隆起后方由面神经分出,经锥隆起内小管到镫骨肌。
从镫骨肌经以下到茎乳孔之间的面神经任一部位分出,经一单独骨管进入并穿过鼓室,然后并入舌神经中。其感觉纤维司舌前2/3的味觉,其副交感纤维达下颌下神经节,节后纤维司下颌下腺与舌下腺的分泌。
分别支配茎突舌骨肌、二腹肌后腹、枕肌、耳后肌、部分耳上肌及耳郭内肌。
面部分支从面神经上(颞面支)、下(颈面支)支再分出5支,支配面部诸肌。上支发出:①颞支,支配额肌、耳前肌、耳上肌、眼轮匝肌及皱眉肌;②颧支,支配上唇方肌与颧肌。下支发出:①颊支,支配口轮匝肌与颊肌;②下颌缘支,支配下唇方肌、降口角肌与颊肌;③颈支,支配颈阔肌。
面神经的内耳道与迷路段主要由迷路动脉的分支供给,乳突段和鼓室段的面神经由茎乳动脉和脑膜中动脉的岩浅支供给。输出静脉主要经茎乳孔和面神经管裂孔到达管外。
声音是由一定的能量作用于可振动的物体所产生,并经某种介质(空气、液体或固体)进行传播的机械振动。在介质中某一质点沿中间轴来回发生振动,并带动周围的质点也发生振动,逐渐向各方向扩展,这就是波,而其中能产生听觉的振动波称为声波。人类能感受声波的频率在20~20 000Hz范围之内。常用的听觉范围,如谈话声仅在500~3 000Hz。频率低于20Hz的声波叫次声波,高于20 000Hz的声波叫超声波。一些动物可听到超声和次声。强度很大时,超声和次声可通过非听觉途径作用于躯体;高于160dB的次声可损伤动物的内脏,导致死亡。声速是声波通过介质传播的速度,它和介质的性质与状态(如温度)有关。介质分子结构越紧密,声速值就越大。声波在空气、水和钢铁中的速度比值约为1∶4∶12。当0℃时,大气中的声速为331m/s;当温度增加到20℃时大气中的声速为343m/s,通常将常温下的声速认定为340m/s。
声音包括两种含义,在物理学上是指声波,在生理学上指声波作用于听觉器官引起的一种主观感觉。尽管这两个含义不同,但他们之间有一定的内在联系。
声音的物理强度是客观的,决定于单位时间内作用于单位面积上能量的大小,可用仪器测量。当一定强度声波作用于人耳后所引起的一种辨别声音强弱的感觉称为响度(loundness)。响度是主观的感受,与声音的物理强度和频率有一定关系。声音必须达到一定强度才能产生听觉,能引起听觉的最小声音强度称为听阈(hearing threshold)。人耳的听阈随着频率不同而各异。在强度相同时,1 000~4 000Hz的声音人耳听起来最响。将各个不同频率的听阈连接成的曲线称为听力图(audiogram)或听力曲线。
频率是声音的物理特性,而音调是频率的主观反映。音调高低与频率的高低一致,但并不成简单的比例关系。频率不变时,音调可因强度不同而稍有差异。当声音在一定强度下,频率与音调的关系相互一致,但当强度增加时,低频率声音的音调显得更低而高频率声音的音调显得更高。
声音主要通过两条途径传入内耳,即空气传导(air conduction)和骨传导(bone conduction)。
简称气导,是声音经外耳道通过鼓膜、听骨链传入内耳的途径。从听觉生理功能看,声波振动被耳郭收集,通过外耳道到达鼓膜,引起鼓膜-听骨链机械振动,进而镫骨足板的振动通过前庭窗使内耳淋巴液产生振动,引起蜗窗基底膜振动,导致基底膜上的螺旋器毛细胞受到刺激产生电活动;毛细胞释放神经递质激动螺旋神经节细胞,产生动作电位。神经冲动沿脑干听觉传导径路到达大脑颞叶听觉皮质中枢而产生听觉。
此外,鼓室内的空气也可先经蜗窗膜振动而产生内耳淋巴压力变化,引起基底膜发生振动。这条径路在正常人是次要的,仅在正常气导经前庭窗径路发生障碍或中断,如鼓膜大穿孔、听骨链破坏中断或固定时才作用明显。
简称骨导,指声波通过颅骨传导到内耳使内耳淋巴液发生相应的振动而引起基底膜振动,耳蜗毛细胞之后的传导过程与气导过程相同。在正常听觉功能中,外界由骨导传入耳蜗的声能甚少,但骨导听觉常用于听力损失的鉴别诊断。
骨导的方式有三种,包括移动式骨导、压缩性骨导和骨-鼓径路骨导。前两种骨导的声波是经颅骨直接传导到内耳,为骨导的主要途径;后一种骨导的声波先经颅骨,再经鼓室才进入内耳,为骨导的次要途径。移动式骨导又称惰性骨导,指声波作用于颅骨时颅骨包括耳蜗作为一个整体反复振动,即移动式振动。压缩式骨导是指声波的振动通过颅骨达耳蜗骨壁时,颅骨、包括耳蜗骨壁随声波的疏密相呈周期性的膨大和压缩。骨-鼓径路骨导是指颅骨在声波作用下振动时,可通过下颌骨髁突或外耳骨壁,将其传至外耳道、鼓室及四周空气中,再引起鼓膜振动。
骨导绕过外耳和中耳直接作用于内耳,因此骨导听阈直接反映耳蜗的功能,这也是临床上应用骨导听阈判断传导性听力损失的原因。但并非骨导听阈只受耳蜗功能的影响,耳蜗外的因素(如改变外耳道压力、堵耳效应、镫骨足板固定、前半规管裂、耳硬化症)也可引起骨导阈值的改变。如耳硬化症可引起2 000Hz骨导阈值明显下降(称Carhart切迹),这可能与镫骨足板固定有关,足板手术后骨导听阈可恢复。
外耳由耳郭和外耳道组成。耳郭可帮助判断声源的方向,其形状有利于收集声波,起采声作用。有些动物能转动耳郭以探测声源的方向,人耳耳郭的运动能力已经退化,但可通过转动头部来判断声源的位置。
外耳道是声波传导的通道,其一端开口于耳郭,另一端终止于鼓膜。根据物理学原理,一端封闭的管道对于波长为其长度4倍的声波能产生最大的共振作用,即增压作用。人类的外耳道长约2.5cm,其共振频率的波长为10cm,按空气中声速340m/s计算,推算人的外耳道共振频率应约为3 400Hz。
声源定位最重要的线索是声波到达两耳时的强度差和时间差。头颅作为障碍物,可产生头影效应,使声音到达左右外耳道口出现耳间强度差(interaural intensity difference,IID)。由于头部的阻挡作用和声波绕射,使声音抵达双耳产生时间差(interaural time difference,ITD)。强度差有利于高频声源的辨向,时间差有利于低频声源的辨向。耳郭还可通过对耳后声源的阻挡和耳前声源的集音而有助于声源定位。因此,头颅、耳郭和外耳道在声源定位中都有重要作用。
中耳的基本功能是将传至外耳道内的声波(机械振动),通过鼓膜的振动和听骨链传递到耳蜗,引起耳蜗内、外淋巴的振动,进而由毛细胞转化为电冲动,经听觉神经系统传递至大脑皮质产生听觉。中耳传递声音的过程类似于一个阻抗匹配器。声波从一种介质传递到另一种介质时透射的能量取决于两种介质声阻抗的比值。阻抗相差越大,声能传递效率越差。由于水的声阻抗远远高于空气的声阻抗,空气与内淋巴的声阻抗相差约3 800倍。当声波由空气传递到淋巴液时,将有99.9%的声能在气液交界面被反射而损失,声能损失约30dB,仅有约0.1%的声能可透射传入淋巴液中。中耳的主要功能是匹配两种传导介质的阻抗差异,避免声音从空气传递到淋巴液引起的声能损失。这种功能主要是由鼓膜与听骨链组成的传音装置完成,即鼓膜与镫骨足板的面积比、锤骨柄与砧骨长突的长度比及鼓膜的喇叭形状产生的杠杆作用。
从声学特性看,鼓膜就像话筒中的振膜,是一个压力感受器,具有良好的频率响应特性和较小的失真度。鼓膜的振动在频率、时程和相位上可完全跟随声波,但振动形式因声音频率不同而有差异。Békésy(1960)应用电容声探头直接研究尸头鼓膜振动时观察到,当频率2 400Hz以下的声波作用于鼓膜时,整个鼓膜沿鼓沟上缘(锤骨前突与砧骨短突的连线)切线的转轴而振动,鼓膜不同部位的振幅大小不同,以锤骨柄向下近鼓膜底部处振幅较大。
人的鼓膜面积约为85mm 2 ,由于鼓膜周边嵌附于鼓沟内,其有效振动面积约为实际面积的2/3,即55mm 2 ,比镫骨足板面积3.2mm 2 大17倍,即声压从鼓膜传至前庭窗膜可增加17倍,相当于声压增加24.6dB。由于鼓膜振幅与锤骨柄振幅之比为2∶1,鼓膜的弧形杠杆作用可使声压提高1倍。
锤骨、砧骨和镫骨以其特殊的连接方式形成杠杆系统。听骨链的运动轴向前通过锤骨柄,向后通过砧骨长脚,支点相当于通过锤骨颈部前韧带与砧骨短脚之间的连线。该杠杆系统的特点是转轴位于听骨链的重心上,在传递能量的过程中惰性最小,效果最好。若加上鼓膜弧度的杠杆作用,则增益更多。声波从空气直接进入内淋巴,因介质阻抗差异而衰减的能量(约30dB),通过中耳的增压作用得到补偿。
鼓膜的振动传至锤骨柄的尖端时,当锤骨柄向内移的瞬间,锤骨头与砧骨体因其在转轴上的位置而向外转;砧骨长突及镫骨因位于转轴的下方,故其运动方向与锤骨柄一致而向内移。Békésy(1951)在人尸体上观察到,在中等强度声压作用时,镫骨足板沿其后脚的垂直轴(短轴)振动。当声强度接近痛阈时,镫骨足板沿其前后轴(长轴)呈摇摆式转动。此时,外淋巴只是在前庭窗附近振动,因而避免了强声引起的基底膜过度位移所造成的内耳损伤。
中耳增压是声波通过鼓膜、听骨链作用于前庭窗时,其振动的压力增大,而振幅减少。中耳的增压放大是通过圆锥形鼓膜的弧形杠杆作用、鼓膜和镫骨足板的面积比(area ratio)及锤骨柄和砧骨长脚的杠杆比(lever ratio)实现的。鼓膜的有效振动面积与镫骨足板面积之比为17∶1,听骨链杠杆系统中锤骨柄与砧骨长脚的长度之比为1.3∶1,故不包括鼓膜弧形杠杆作用在内的中耳增压效果为22.1倍,相当于27dB。声波从空气直接进入内淋巴,因声阻抗不同而衰减的能量,通过中耳的增压作用得到部分补偿。
正常情况下,声音经鼓膜、听骨链和镫骨足板作用于前庭窗,振动耳蜗外淋巴,再经前庭阶、蜗孔、鼓阶,最后到达蜗窗膜。由于声音传导途径的差异使到达两窗的声波相位不同,作用于两窗上的声音振动不相互抵消,为声波在外淋巴中的传导提供有利条件。若蜗窗病变(如固定),虽然前庭窗仍活动,但不能推动外淋巴振动,可出现传导性听力损失。研究表明,10%鼓膜穿孔面积可导致2 000Hz以下频率约15dB的听阈提高,但2 000Hz以上频率听力损失较少;鼓膜缺如时可造成2 000Hz以下频率约45dB的听阈提高,而在3 000Hz处听力损失可达50dB。可见听力损失随着穿孔面积的增大而增加。这可能是穿孔使鼓膜有效振动面积与镫骨足板面积之比率减少所致。鼓膜穿孔对声波不同频率的影响。当整个鼓膜大穿孔(鼓膜缺如)时听力损失程度大于中耳增压作用的30dB,是由于声波可同时到达前庭窗和蜗窗,使两窗之间的压差抵消之故。
中耳肌肉包括鼓膜张肌和镫骨肌。前者由三叉神经下颌支支配,后者由面神经镫骨肌支支配。鼓膜张肌收缩时将锤骨柄与鼓膜向内牵引,使鼓膜的紧张度增加,各听小骨之间连接更为紧密,并引起镫骨足板推向前庭窗,使内耳外淋巴压力增高。镫骨肌收缩时牵引镫骨头向后移动,使足板前部向外翘起,导致内耳外淋巴压力减少。二者相互作用,可降低中耳传音功能。
在受外界声音和非声音刺激时,中耳肌肉可反射性收缩。声音刺激如纯音和白噪声;非声音刺激包括电、气流刺激及触碰外耳道等。由于鼓膜张肌对机械性刺激敏感,而对声音刺激的阈值高于镫骨肌反射阈。因此,在声音刺激引起人类的中耳肌反射收缩中镫骨肌收缩起主要作用。
当鼓膜张肌和镫骨肌同时收缩时,鼓膜内移,听骨链被压缩,中耳劲度阻抗明显增加,可使1 500Hz以下的声音衰减10dB左右。中耳肌反射对高强度声刺激的保护作用,低频较高频明显。但因该反射有一定潜伏期,对于突发性的爆震声保护作用不大。
正常情况下,由于近鼻咽腔管道的软骨管壁的弹性作用和周围组织的压力及咽部的牵拉作用,咽鼓管的咽口经常处于闭合状态。当吞咽、打哈欠、打喷嚏以及咀嚼等动作时,通过腭帆张肌、腭帆提肌及咽鼓管咽肌(其中腭帆张肌起主要作用)的收缩,咽鼓管瞬间开放,调节鼓室内气压,使之与大气压保持平衡。鼓室两侧压力相等,使中耳阻抗保持最低值,有利于鼓膜的振动和听骨链的传导。鼓室与外界气压差达到一定数值时,也可引起咽鼓管开放。当鼓室内气压大于外界气压时,气体通过咽鼓管向外排放比较容易;而当外界气体大于鼓室内气压时,气体的进入则比较困难。
咽鼓管黏膜上皮细胞表面有丰富的纤毛,鼓室黏膜及咽鼓管黏膜的杯状细胞与黏液腺所产生的黏液借助纤毛运动,不断向鼻咽部流出。
正常情况下,咽鼓管处于闭合状态,可阻挡嗓音、呼吸、心跳等自体声音经鼻咽部、咽鼓管直接进入鼓室。咽鼓管异常开放的患者,说话时咽鼓管不能处于闭合状态,声波经开放的咽鼓管直接进入中耳腔,引起鼓膜振动过强而产生自听过响(autophonia)症状。呼吸时引起的空气流动也可通过开放的咽鼓管进入中耳腔产生呼吸声,这种呼吸声可掩蔽经外耳道传入的外界声响。由于咽鼓管外1/3段(咽鼓管骨部)通常处于开放状态,呈逐渐向内(软骨部)变窄的漏斗形,且表面被覆部分皱褶的黏膜,类似吸音结构,可吸收因蜗窗膜及鼓膜振动而引起的鼓室腔内的声波,故有消声作用。
正常人咽鼓管平时处于闭合状态,仅在吞咽瞬间才开放,来自鼻腔的温暖、湿润的气体在咽鼓管开放的瞬间进入中耳。咽鼓管软骨部黏膜较厚,黏膜下层有疏松结缔组织,使黏膜表面产生皱襞,后者具有活瓣作用,加上黏膜上皮的纤毛运动,对阻止鼻咽部的液体、异物及感染灶等进入鼓室具有重要意义。
耳蜗的功能可概述为:①感音功能,即将传入的声能转换成适合刺激蜗神经末梢的形式;②对声音信息的编码,即分析传入声音的特性(频率与强度),使大脑能处理该刺激声中包含的信息。
当声音作用于鼓膜时,声波的机械振动通过听骨链传递到前庭窗,压力变化立即传给耳蜗内的液体和膜性结构。当前庭窗膜内移,前庭膜和基底膜也将下移,最后鼓阶的外淋巴压迫蜗窗膜,使蜗窗膜外移;当前庭窗膜外移时,整个耳蜗内的液体和膜性结构又向反方向移动,如此反复,形成振动。振动从基底膜的底部开始,按照物理学中的行波(traveling wave)原理向耳蜗的顶部方向传播。
由于耳蜗底部的基底膜劲度最大,能量从耳蜗内液体传至该部分的效率最大。另外基底膜底部距离前庭窗膜和蜗窗膜最近,根据能量传递的就近原则,声波传入内耳后将首先通过底部基底膜向蜗窗传播。因此耳蜗底部的基底膜首先振动,再以行波的形式向顶部传播。振动在基底膜上从耳蜗底部向顶部传播时,振幅逐渐增大,而传播速度逐渐变慢,波长变短。当振动达到基底膜某一部位,即基底膜共振频率与声波频率一致时,振幅最大;离开该处后,振幅迅速减小,在稍远处基底膜的位移完全停止。声波频率越高,行波传播越近,最大振幅出现的部位越靠近前庭窗处;声波频率愈低,行波传播的距离愈远,最大振幅出现的部位愈靠近蜗顶。行波的速度在行波向耳蜗顶部移行的过程中逐渐减慢,故行波的相位随着传导距离的增加而改变,其波长亦逐渐减少,但在蜗管任何点的振动频率都与刺激声波的频率相同。
耳蜗对声音的分析编码功能和一般感受器相同,即不同频率的声音通过基底膜不同部位的毛细胞兴奋,经过不同的神经纤维传入中枢的特定部位形成。基底膜上不同位置的毛细胞,由于他们的机械特性、耦合特性和电共振特性不同,都有自己的特征频率。特征频率的声音只需极低的强度,就能引发该部位毛细胞反应,而低于或高于特征频率的声音需要很高强度才能引发该毛细胞同样的反应。不同频率的声音引起毛细胞反应所需强度的曲线,称为该毛细胞的谐振曲线(tuning curve)。此外,听神经发放不同频率的冲动来传递声音频率信息。频率低于400Hz的声音,听神经大体按声音的频率发放冲动。400~5 000Hz声音作用时,神经纤维分成若干组,每组纤维间隔若干声波周期发放一次冲动,互相错开,以此进行,各组纤维同时发放的总数与声音频率接近。耳蜗对信息频率的编码就是通过上述的部位原则和频率原则实现的。
高强度声音通过基底膜振动的幅度和速率增加,使中枢感觉到声音增强。当声音强度增大时,基底膜振动的振幅变大,毛细胞感受器电位增大,使神经纤维发放动作电位频率增加,中枢感觉到声音变响。声音强度增加还可使基底膜振动的速率增加,被兴奋的毛细胞数目增多,复合听神经动作电位的幅度变大,中枢感觉到响度增强。由于基底膜振动的幅度和速率增加,将兴奋某些特殊的内毛细胞,传入冲动到中枢,感觉到声音响亮。
哺乳动物耳蜗对声音刺激的反应具有非线性特点。耳蜗基底膜机械反应的强度与传入声波的强度具有压缩式或扩展式非线性特点。这与外毛细胞的主动收缩有关,它使人耳听觉的强度范围变得很大,也使人耳在听到强声时不会感到特别强烈。耳蜗的许多非线性现象与两音干涉有关,如双音抑制(two-tone suppression)和相互调制畸变(intermodulation distortion)。听神经纤维对单个纯音的刺激仅表现为兴奋性反应,没有抑制效应。然而一个纯音存在可影响听神经纤维对另一个纯音刺激的反应。如果适当安排某两种纯音的频率和强度,则第二种纯音能抑制或压制听神经纤维对第一种纯音的刺激反应,该现象称为双音抑制。相互调制畸变指当同时给予频率不同的两个或多个初始音刺激时,人耳可听到不同于刺激声频率的另外频率的声音,这是因为初始音的相互调制产生了畸变产物,即与初始音有特定频率关系的另外频率的声音。
蜗内电位(endocochlear potential,EP)又称内淋巴电位,系蜗管内淋巴与鼓阶淋巴之间的电位差所致。该电位差是所有耳蜗生物电反应产生的基础,也是耳蜗完成声-电转换功能的基础。EP起源于血管纹,有助于听觉感受器将声能转变为神经冲动。缺氧或代谢抑制时,能使EP迅速下降。
耳蜗微音器电位(cochlear microphonics,CM)是耳蜗受声音刺激时,在耳蜗及其附近结构所记录到的一种与声波的频率和幅度完全一致的交流性质的电位变化。在听阈范围内,CM能重复声波的频率,主要产生于外毛细胞。
总和电位(summating potential,SP)是指耳蜗受声音刺激时,毛细胞产生的一种直流性质的电位变化,产生于内毛细胞。
听神经动作电位(action potential,AP)是指耳蜗对声音刺激产生的蜗神经末梢的动作电位,具有传递声音信息的作用。
听觉中枢结构包括蜗神经核、上橄榄核、斜方体核、外侧丘系核、下丘、内侧膝状体及听皮层等。听觉传入通路的第一级神经元为耳蜗听神经元(又称耳蜗螺旋神经节细胞),位于耳蜗的骨性骨螺旋小骨内。人类约有30 000个听神经元,其中约有95%为Ⅰ型神经元,其周围突与内毛细胞构成突触连接;约5%为Ⅱ型神经元,与其周围突与外毛细胞构成突触连接。第二级神经元的胞体位于耳蜗腹侧核和耳蜗背侧核,发出的纤维大部分在脑桥内斜方体,并交叉至对侧,至上橄榄核外侧折向上行,组成外侧丘系。外侧丘系的纤维经中脑被盖的背外侧部,大多数终止于下丘。第三级神经元的胞体位于下丘,其纤维经下丘臂止于内侧膝状体。后者发出纤维组成听辐射,经内囊到达大脑皮质颞横回的听区。
听觉传导通路在蜗神经核以上各级中枢都接受双侧耳传来的信息,因此若一侧通路在外侧丘系以上受损,不会产生明显的症状;但若损伤了蜗神经、内耳或中耳,则将导致听觉障碍。
听觉的发射中枢在下丘。听觉的传入冲动到达下丘后,下丘神经元发出纤维到达上丘,后者发出的纤维经顶盖脊髓束下行至脊髓,控制脊髓前角细胞的活动,完成听觉反射。
此外,听觉通路具有较强的反馈作用。例如,大脑皮质听区还可发出下行纤维,经听觉通路上的各级神经元中继,影响内耳螺旋器的感受功能,形成听觉通路的反馈调节。
人体经常感受线性加速和角加速运动,运动中人的体位和周围环境都会发生改变,需要通过前庭、视觉和本体感觉三个系统组成的“平衡三联”维持身体平衡,其中前庭系统是专司平衡的器官,由前庭感受器、前庭神经、前庭神经核(vestibular nuclei,VN)、若干神经传导通路和3级调控中枢构成(图2-3-1)。
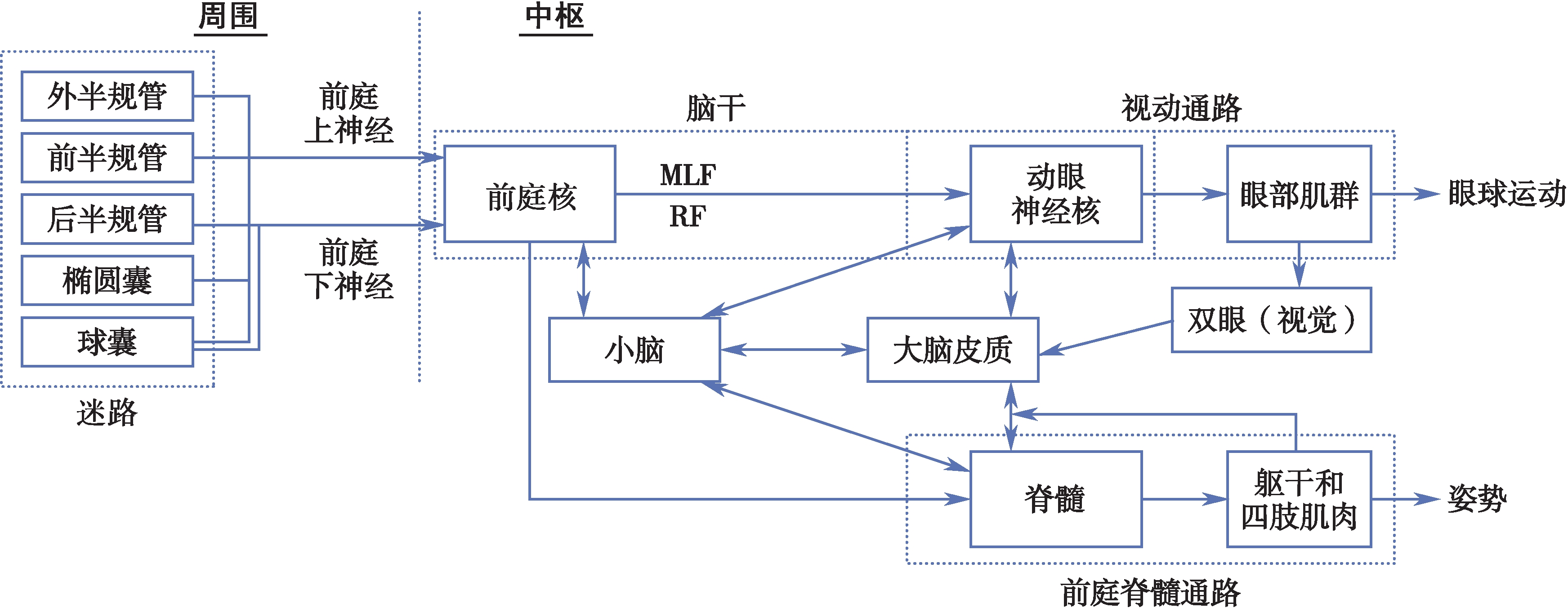
图2-3-1 前庭系统组成示意图
前庭感受器包括3对半规管(semicircular canal)和2对耳石器(otolith organ),前者指外半规管(lateral semicircular canal,LSCC)、前半规管(anterior semicircular canal,ASCC)和后半规管(posterior semicircular canal,PSCC);后者指椭圆囊(utricle)、球囊(saccule)。前庭(vestibule)、半规管和耳蜗一起组成骨迷路(bony labyrinth)。
前庭神经起源于内耳前庭神经节(Scarpa's ganglion)的双极细胞,分为上、下两部分,由神经分支相联系,接受来自半规管和耳石器的传入信号,并将其继续传入前庭神经核。
前庭神经节为前庭系初级神经元,前庭神经核为前庭系次级神经元。迷路和前庭神经组成外周前庭系统(peripheral vestibular system)。
前庭神经核是前庭系的初级信息整合中心,通过眼动通路(oculomotor pathway)、前庭-脊髓通路(vestibulospinal pathway)、前庭-小脑通路等,与动眼神经核、脊髓前角运动神经元及小脑相连,产生前庭-眼反射(vestibulo-ocular reflex,VOR)、前庭-脊髓反射(vestibulo-spinal reflex)等,保持视觉清晰、调整姿势,保持平衡。前庭神经核还与脑干网状结构、自主神经系统、大脑皮质等有广泛联系,感知自身空间位置、保持自主神经系统功能正常等。
前庭系统功能由脑干、小脑、大脑皮质3级中枢进行调控。
前庭位于耳蜗及半规管之间,略呈椭圆形,前下部较窄,有一椭圆孔通入耳蜗的前庭阶;后上部较宽,有3个骨半规管的5个开口通入。其外壁即为鼓室内壁,上有前庭窗及蜗窗;内壁为内耳道底;上壁骨质中有面神经迷路段穿过,前庭腔内面有从前上方向后下方弯曲的斜形骨嵴,称前庭嵴(vestibular recess)。嵴的前方为球囊隐窝(spherical recess),内含球囊。嵴的后方有椭圆囊隐窝(elliptical recess),内含椭圆囊(图2-3-2、图2-3-3)。
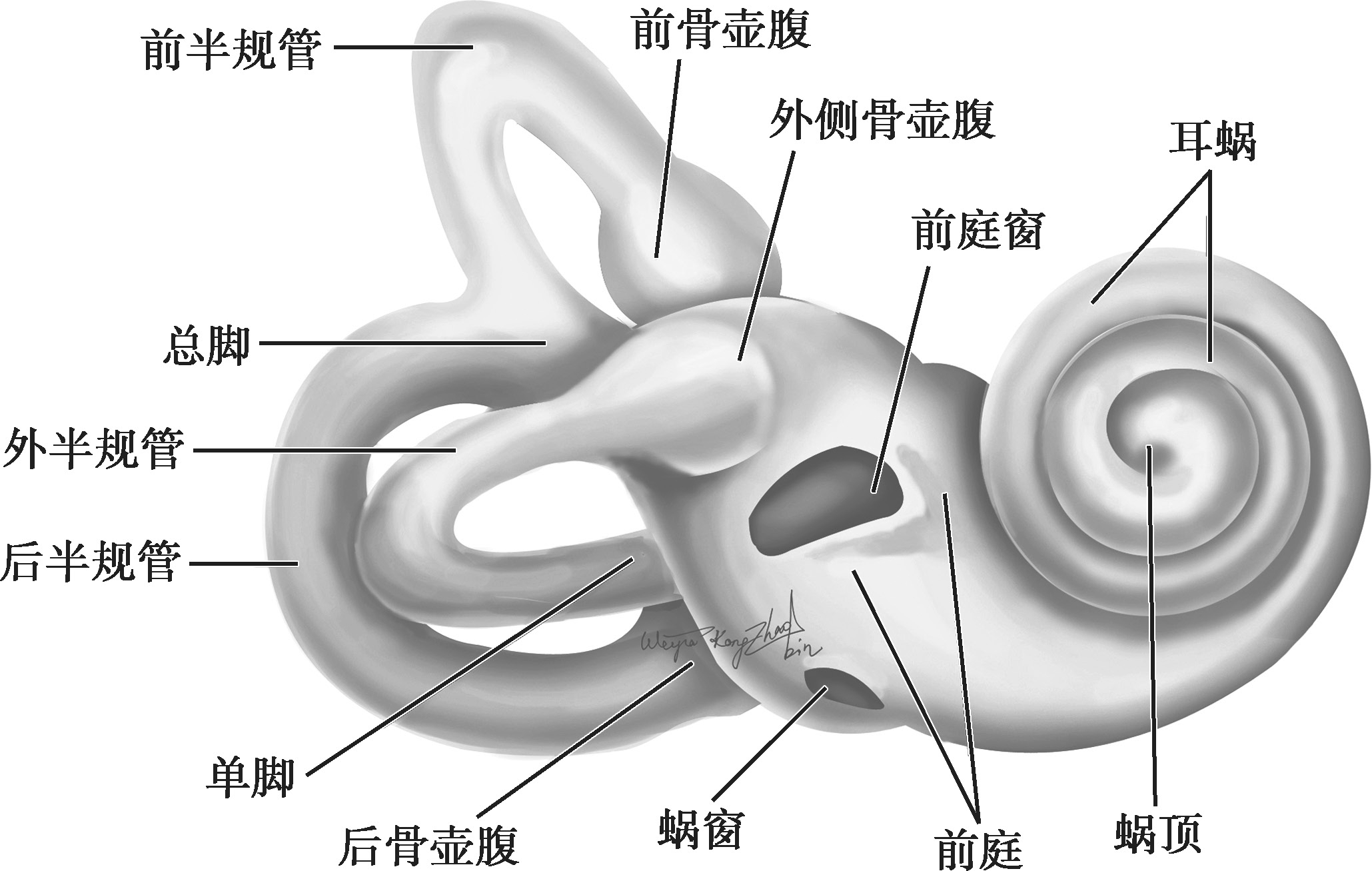
图2-3-2 右侧骨迷路(外面观)
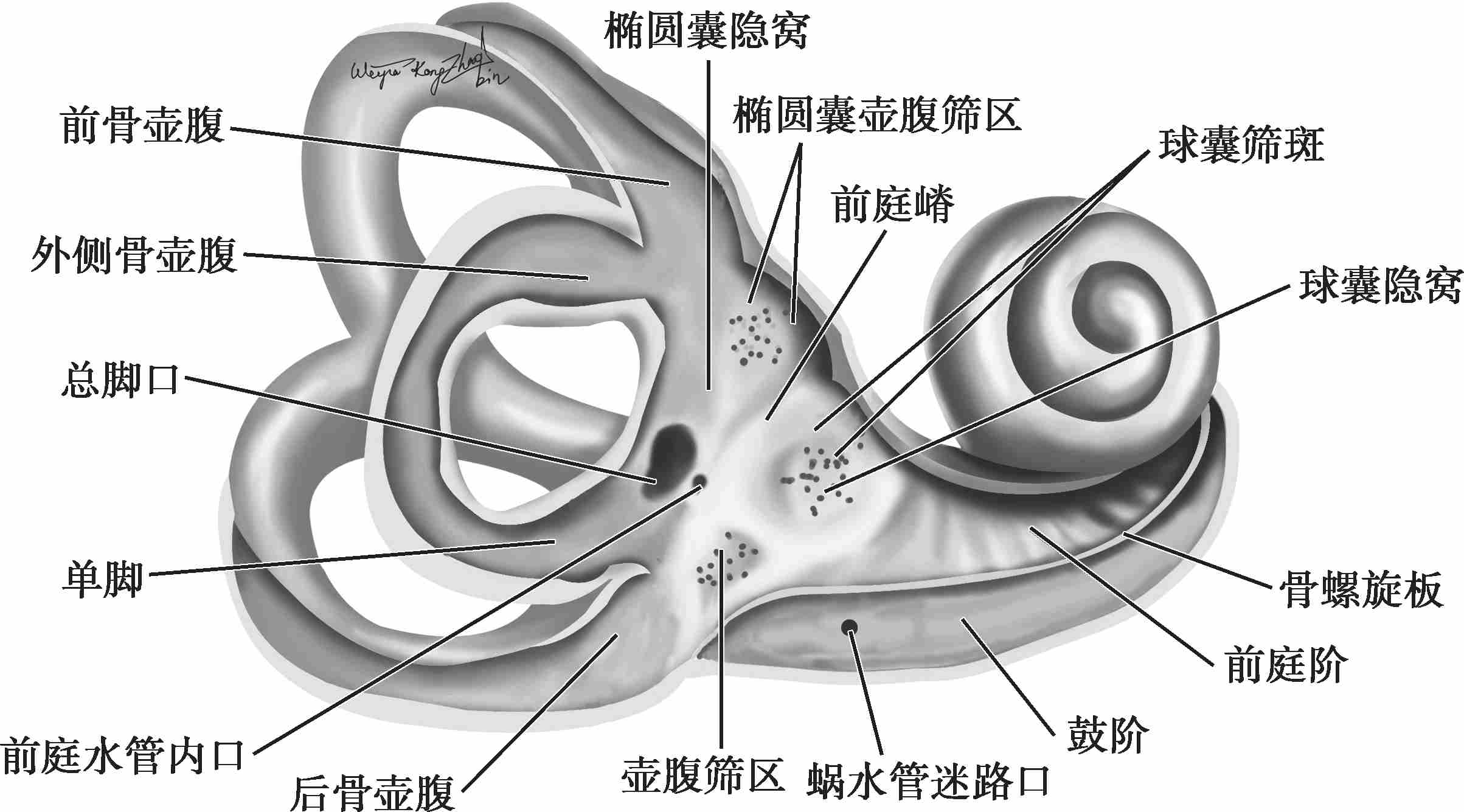
图2-3-3 右侧骨迷路(半规管、前庭、耳蜗底转被揭开)
半规管分为骨半规管(osseous semicircular canals)和膜半规管(membranous semicircular canals)。
骨半规管为3个2/3环形的小骨管,依其所在的位置,分别称为外半规管、前半规管和后半规管。外半规管前部与Reid基线(眶下缘至外耳门中点的连线)向上大约30°(图2-3-4A)。
双耳共有6个半规管,依据其感受平面分为3组(图2-3-4B):①双侧外半规管为一组,当头前倾30°时,外半规管平面与地面平行;②左侧前半规管和右侧后半规管为一组(left A SCC right P SCC,LARP);③右侧前半规管和左侧后半规管为一组(right A SCC left P SCC,RALP)。
三个半规管平面之间大约成90°角,但准确地说后半规管与外半规管平面成92°角,而前半规管与外半规管平面成90°角。外-后半规管平面的夹角为75.8°~98.0°,外-前半规管平面的夹角为77.0°~98.4°,后-前半规管平面的夹角为75.8°~100.1°。因此,当头部转动时,每个半规管都有可能同时感受到来自不同角度的刺激。因此临床上如果良性位置性眩晕(benign positional vertigo)手法复位效果不佳时,可以考虑适当偏移方向后再行复位。
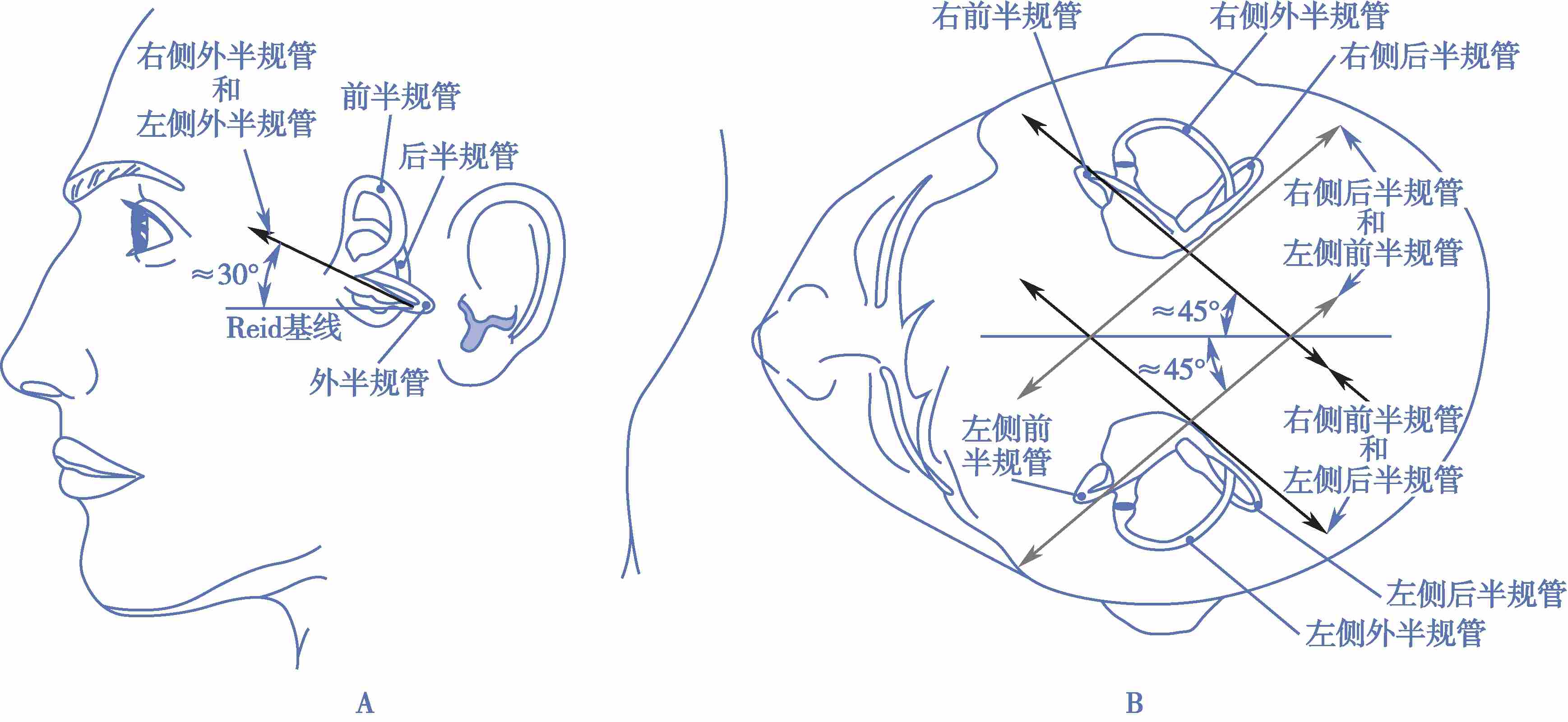
图2-3-4 半规管位置示意图
A.黑色箭头所示为左侧外半规管与Reid基线夹角;B.图中两条黑色平行线所示为RALP平面,深灰色平行线标记为LARP平面。
膜半规管借纤维束固定于骨半规管内,悬浮于外淋巴(perilymph)中,借5个开口与椭圆囊相通,约占骨半规管内腔的1/4,但膜壶腹几乎充满骨壶腹的大部空间。膜半规管内充满内淋巴(endolymph),内淋巴比重略高于水,成分与细胞内液相似,高K + 而低Na + ,而外淋巴成分类似于细胞外液。
半规管末端膨大形成壶腹(ampulla),内有胶样杯状组织称为嵴顶(cupula terminalis)或嵴帽。嵴帽是半规管和前庭之间的屏障,沿半规管内腔延伸,完全覆盖在壶腹的横截面上,其顶端附有黏多糖,感受头部位置的动态改变。
嵴帽的下方是壶腹嵴(crista ampullaris),呈鞍形,垂直于半规管。壶腹嵴上有高度分化的感觉上皮,有前庭神经壶腹支分布,是重要的平衡感受器,由毛细胞和支持细胞组成(图2-3-5)。
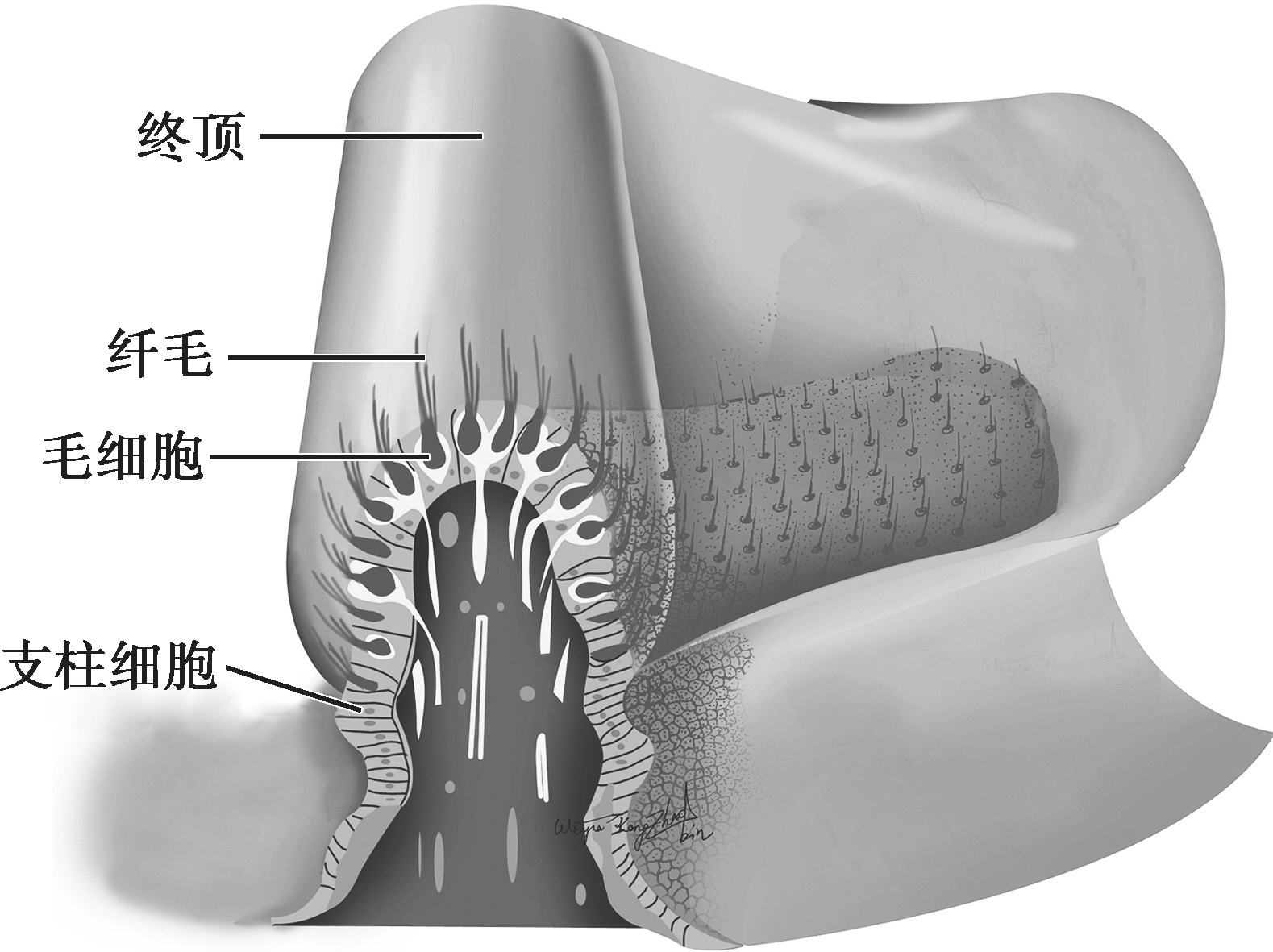
图2-3-5 壶腹嵴模式图
前庭毛细胞分两种类型(图2-3-6):Ⅰ型毛细胞呈烧瓶状,胞体被神经盏(nerve chalice)样传入神经末梢所包绕,细胞之间以1∶1或2∶4的比例形成突触联系;Ⅱ型毛细胞呈柱状,细胞之间形成多突触联系。
每个毛细胞顶端有1根动纤毛和50~100根静纤毛。动纤毛位于一侧边缘,最长,较易弯曲;静纤毛以动纤毛为排头,按从长到短排列,离动纤毛越远则长度越短。当静纤毛束向动纤毛弯曲时,毛细胞去极化(兴奋),反之则超极化(抑制)。嵴帽下端有直径3~5mm的开口,动纤毛和静纤毛束由此伸入嵴帽。
椭圆囊底部的前外侧有椭圆形、较厚的感觉上皮区,即椭圆囊斑(macula utriculi),分布有前庭神经椭圆囊支。后壁有5孔,与3个半规管相通。前壁内侧有椭圆囊管,连接球囊与内淋巴。
球囊较椭圆囊小,其前壁有球囊斑(macula sacculi),呈匙状,分布有前庭神经球囊支。椭圆囊斑和球囊斑互相垂直,均由毛细胞和支持细胞组成(图2-3-6)。毛细胞纤毛上方覆有一层胶体膜,即耳石膜(otolith membrane),由多层以碳酸钙结晶为主的颗粒即耳石(otolith)和蛋白凝合而成,耳石可以为耳石器的运动提供惯性,加重内淋巴移动时的重力作用。
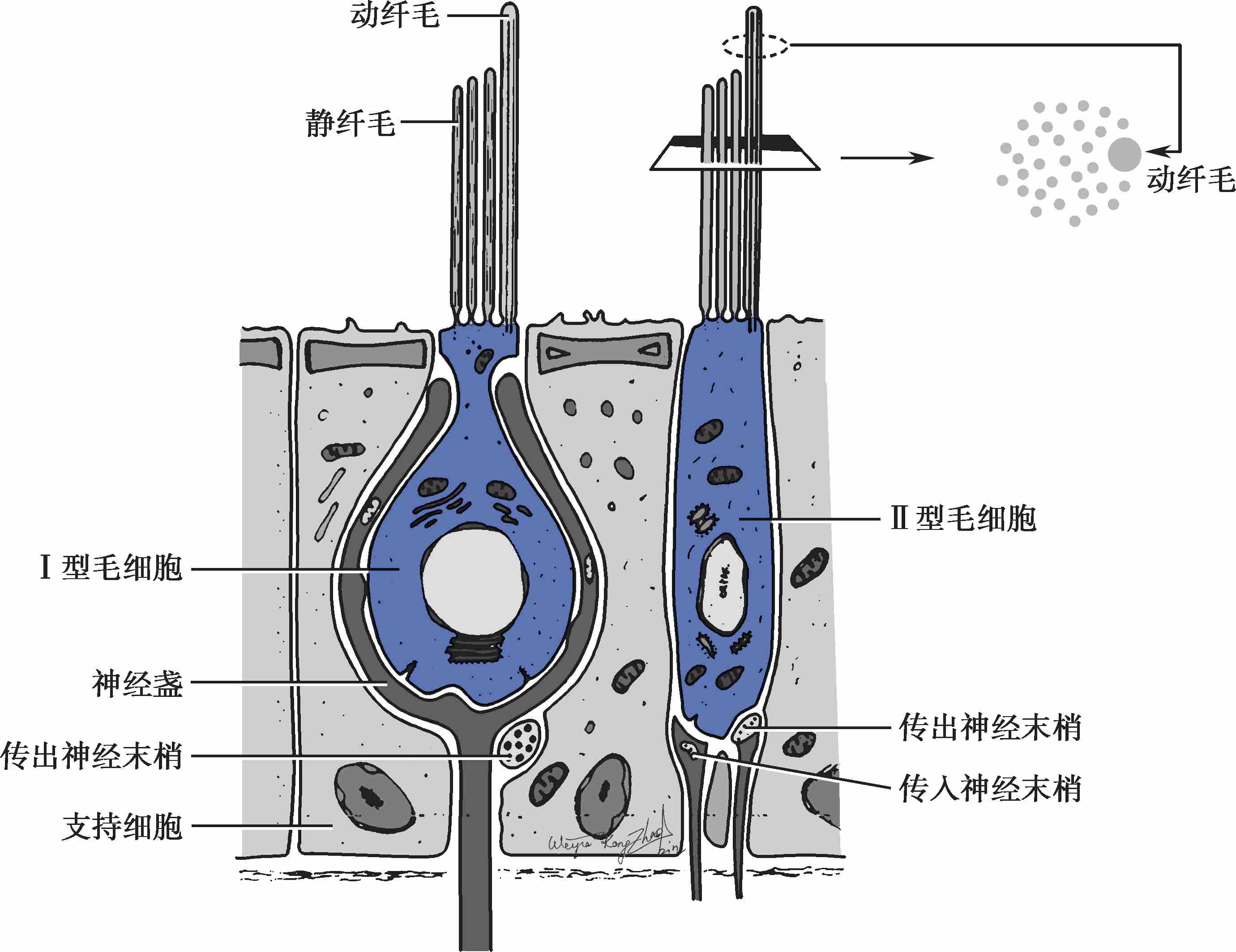
图2-3-6 前庭Ⅰ型、Ⅱ型毛细胞模式图
囊斑表面中央部分有微纹(striola)(图2-3-7),椭圆囊斑微纹呈U形,球囊斑微纹呈L形。微纹将耳石器一分为二,两侧毛细胞上动纤毛的极性相反:椭圆囊斑动纤毛弯曲方向朝向微纹,而球囊斑动纤毛弯曲方向则是背离微纹。这种结构特点使耳石器可感受到来自各方向的线性加速度刺激,并且其中总有一部分呈兴奋状态,而另一部分呈抑制状态,因此耳石器的每个部分在对侧都有各自相对应的功能配对区域。
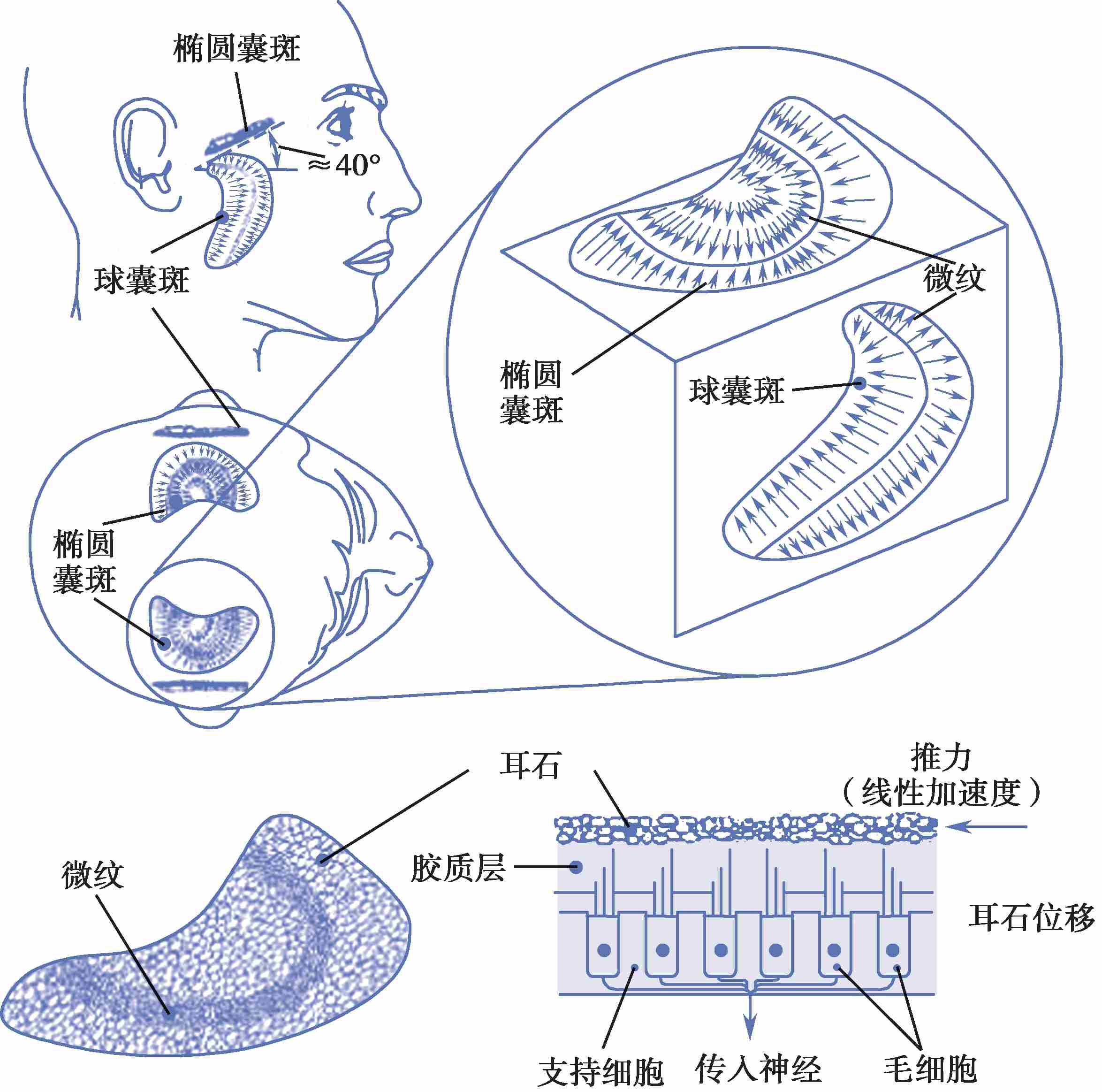
图2-3-7 囊斑示意图
前庭上神经(superior vestibular nerve)穿过内耳道底之前庭上区的小孔,分布于前半规管壶腹嵴(前壶腹神经)、外半规管壶腹嵴(外壶腹神经)、椭圆囊斑(椭圆囊神经),另有分支分布于球囊斑前上部(Voit神经)(图2-3-8)。
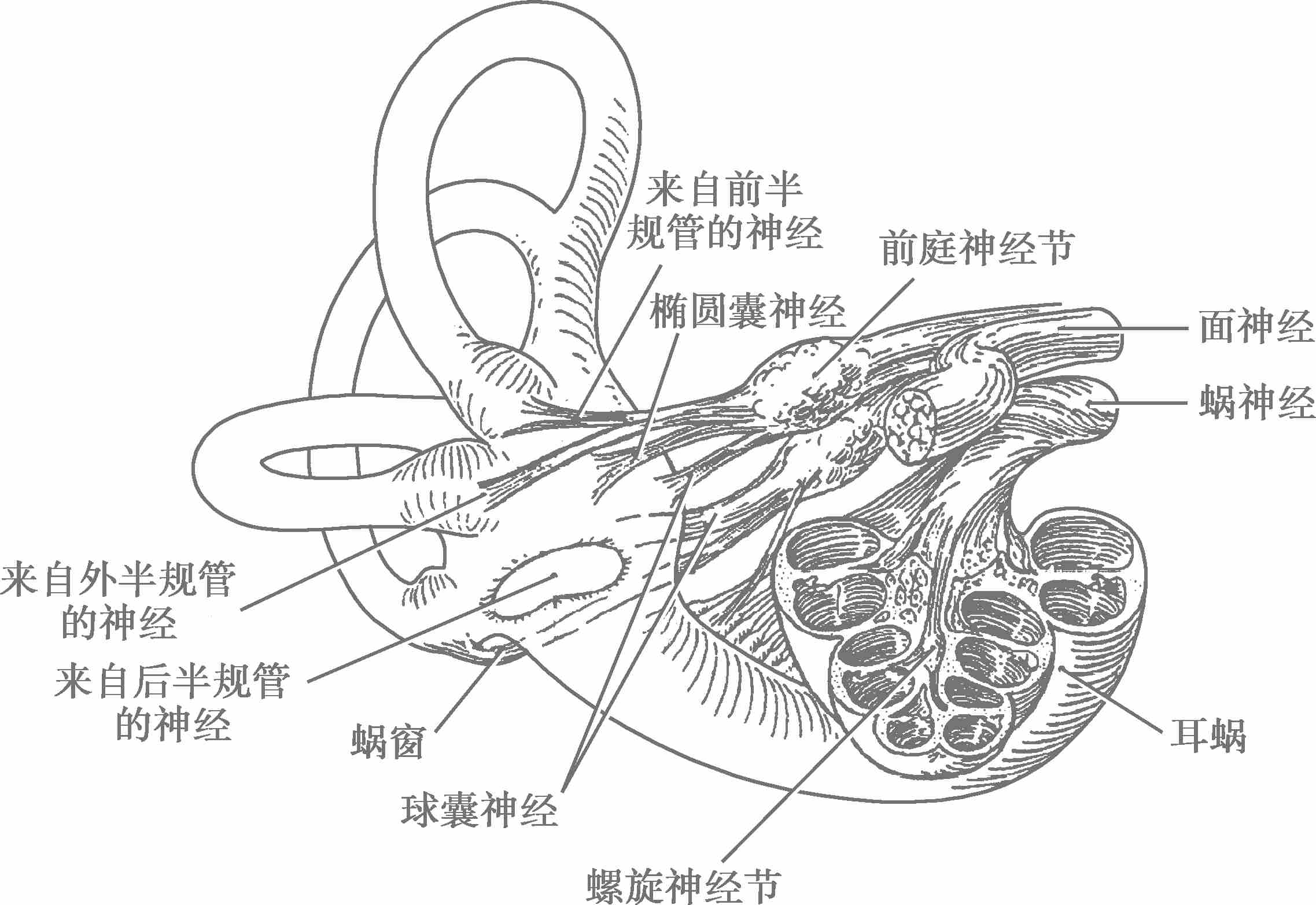
图2-3-8 前庭神经分布示意图
前庭下神经(inferior vestibular nerve)穿过内耳道之前庭下区,分布于球囊斑(球囊神经)、后半规管壶腹嵴(后壶腹神经)。
迷路血供来自迷路动脉,后者主要来自前下交通动脉(anterior inferior cerebellar artery),还有部分来自小脑上动脉或基底动脉。迷路动脉发出分支,其中前庭上动脉(anterior vestibular artery)供应前半规管、外半规管和椭圆囊,以及小部分球囊;前庭后动脉(posterior vestibular artery)供应后半规管和大部分球囊。静脉回流与动脉伴行。前庭上静脉接受来自前半规管、外半规管和椭圆囊的血液回流,前庭后静脉接受来自球囊、后半规管的血液回流。
前庭毛细胞在静息状态下放电频率大约为70~100次/s,Ⅰ型毛细胞的放电是不规则的,而Ⅱ型毛细胞的放电是规则的。所以,两类毛细胞分别感知不同频率和加速度的头部运动,前庭中枢的传入也分不规则性和规则性两种:①Ⅰ型毛细胞的不规则性输入对头部高加速度转动更敏感,帮助中枢在VOR启动时即可迅速探得头部运动。此外,在角前庭-眼反射(angular vestibuloocular reflex,aVOR)中,Ⅰ型毛细胞还参与察觉与距离有关的变化。但是,不规则性输入对头部的低频和较小加速度时产生的VOR没有作用。②Ⅱ型毛细胞的规则性输入,可以在更大的频率范围内,向前庭中枢提供与头动速度成比例的输入信号。
头部向某一侧转动时,内淋巴随之流动,在嵴帽两侧造成的压力差,使前庭毛细胞的静纤毛向动纤毛弯曲,同侧毛细胞兴奋,同时对侧毛细胞被抑制。兴奋时,初级传入神经元放电频率可由静息时的70~100次/s增加到400次/s;而对侧神经元被抑制时,放电频率降低到0次/s后就不再下降,即抑制性中断(inhibitory cutoff)。因此双侧神经元的兴奋和抑制程度是不等的,即兴奋大于抑制,这是Ewald第二、第三定律的生理基础,为半规管和耳石器的共同特性,也是视频头脉冲试验(video head impulse test,v-HIT)和摇头试验(head-shake test)的生理基础。双侧周围前庭传入的兴奋和抑制程度的不一致性,触发前庭中枢的平衡调节机制,使身体达到新的平衡状态。
Ewald(1892)提出了半规管平面、内淋巴流动方向与诱发性眼震、头部运动方向之间的关系,称为Ewald定律(Ewald's Law)。
半规管受刺激时,在该半规管作用平面产生与头动有关的眼球运动。
在水平向的半规管,内淋巴向壶腹流动时引起较强的反应(眼震或头部运动),而内淋巴离壶腹流动时引起较弱的反应,反应强度比为2∶1。
在垂直向的半规管,内淋巴离壶腹流动时引起较强的反应,而内淋巴向壶腹流动引起较弱的反应。
半规管主要感受头部旋转运动,即角加速度改变。当半规管在角加速度的作用下旋转时,内淋巴由于惰性,其流动方向与旋转方向相反。一旦内淋巴开始流动,嵴帽就会封闭其流动的通道,以阻止其流动。此时,由于内淋巴流动造成的压力差,使嵴帽中央部分呈波浪翻滚样的变形运动,壶腹嵴的基底部和顶部也会产生轻度运动。嵴帽的中央区域变形最大,所以此处壶腹嵴表面产生的剪切力也最大,导致静纤毛束运动,使毛细胞转换通道开放(或关闭),细胞膜静息电位随之发生改变:当静纤毛朝向动纤毛方向弯曲时,毛细胞兴奋(去极化),膜电位改变,称启动电位,引起毛细胞释放神经递质,作用于传入神经末梢,此为毛细胞的机械-电能转换。反之,当静纤毛背离动纤毛方向弯曲时,毛细胞抑制(超极化)。
根据Ewald第二、第三定律,在水平向的半规管(如外半规管),内淋巴向壶腹(椭圆囊)移动时,毛细胞兴奋;在垂直向的半规管(如前半规管和后半规管),内淋巴离壶腹(椭圆囊)移动时,毛细胞兴奋。三组半规管在各自的共同作用平面呈推拉(push-pull)的动力学改变。例如,当头部右转时,右侧外半规管毛细胞兴奋,而左侧外半规管抑制。大脑通过比较双侧外半规管的输入差异,判定头部运动的方向。
椭圆囊斑感受水平向、球囊感受垂直向的线性加速度改变。
头部转动时,产生线性加速度,刺激耳石器,反射性地产生眼动,其方向和头部转动方向相反,以保持视觉清晰。重力也可以看成一种向下的线性加速度。
在线性加速度的作用下,由于耳石膜中耳石的比重大于内淋巴,因此耳石的运动略滞后于内淋巴流动(惰性),导致耳石膜向逆作用力方向发生位移,在耳石膜与囊斑毛细胞表面产生剪切力,使毛细胞静纤毛弯曲,启动毛细胞的机械-电能转换过程,通过传入神经传向前庭神经核。当静纤毛向动纤毛方向弯曲时,囊斑兴奋,反之则抑制。
囊斑毛细胞可感知头位变动,再通过前庭核、内侧纵束、动眼神经核,在锥体外的控制下,使眼球反向运动,使目标仍然落在视网膜黄斑部位,维持视野清晰。
双侧囊斑感受各方向的线性加速度改变,调整四肢肌张力,进而调整身体姿势和体位,保持平衡。其中,球囊斑主要调整四肢内收和外展肌的张力,而椭圆囊斑主要调整躯体伸肌和屈肌的张力。
前庭神经核复合体位于脑干延髓背侧,由前庭上核、前庭下核、前庭内侧核和前庭外侧核组成。前庭内侧核、前庭上核位于前庭核腹部,主要接受来自半规管的信号,向中枢投射,主要与角VOR(angular VOR,aVOR)的产生和前庭补偿有关。前庭下核和外侧核位于前庭核尾部,接受来自耳石器的刺激,与线性VOR(translational VOR,tVOR)的产生有关,并作用于前庭脊髓反射。
前庭神经核的传入有部分直接来自前庭神经,另外前庭神经核还接受来自颈部、脊髓、小脑、网状结构以及对侧前庭神经核的传入(图2-3-9)。
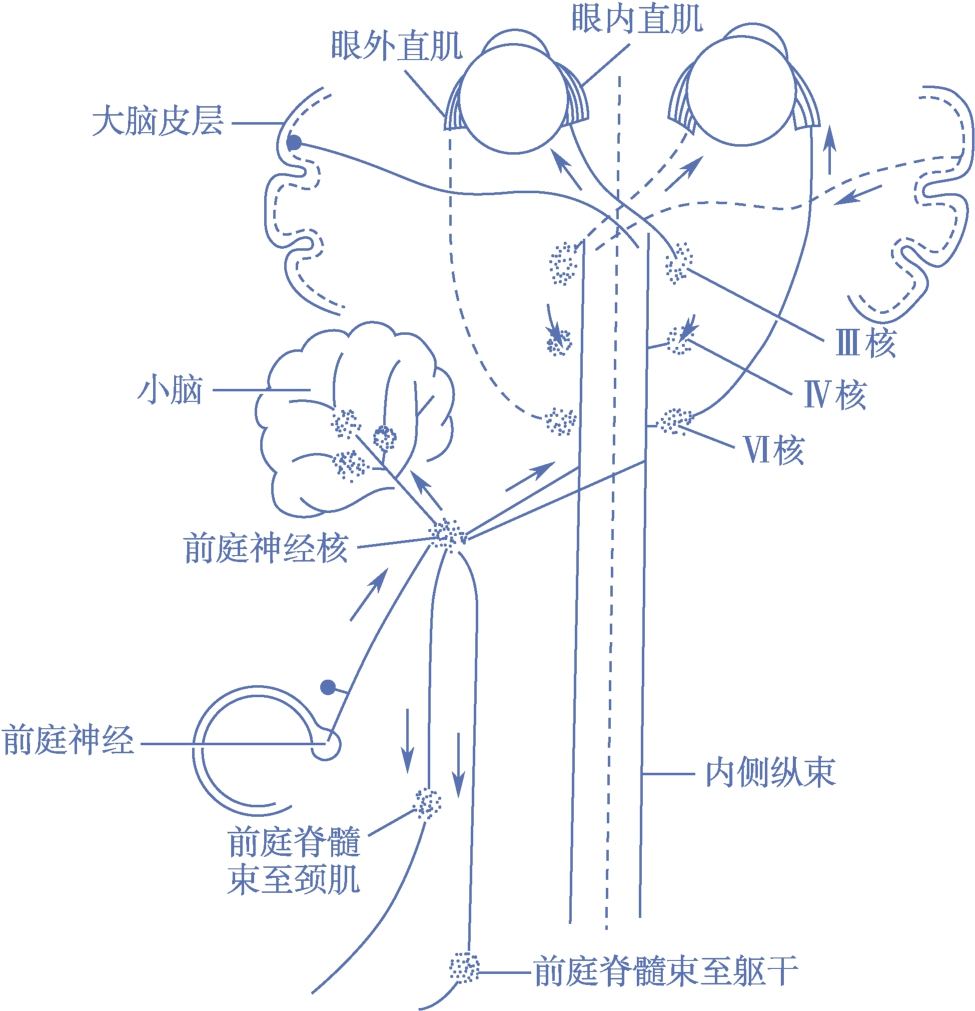
图2-3-9 前庭神经传导径路
前庭神经核传出通路较为复杂,归纳如下(见图2-3-1,图2-3-9)。
经内侧纵束(medial longitude fasciculus,MLF),到达展神经核、动眼神经核、滑车神经核等控制眼动的核团。
通过前庭脊髓外侧束、前庭脊髓内侧束、前庭脊髓尾束和网状脊髓束,到达脊髓前角细胞运动神经元。
小脑和前庭神经核、动眼神经核、大脑皮质(顶叶和岛叶)、脊髓之间存在双向联系;前庭神经的传入信号也可以直接到达小脑。传入信息经小脑下脚绳状体(juxtarestiform body),到达前庭小脑(绒球小结叶)。此外,小脑接受从脑桥核、网状核、旁正中束和下橄榄核等区域传入的视觉和眼动信号。因此,虽然小脑不是VOR反射弧的一部分,却是VOR增益(gain)调节的关键部位。VOR增益是指眼动和头动之间的比率,增益调节发生于单侧前庭功能损伤或服用某些前庭抑制剂时。
中枢前庭系统和网状结构、自主神经系统、下丘、大脑皮质有密切联系,因此刺激前庭会出现眩晕、眼震、平衡失调、倾倒、自主神经反应、空间定向障碍等临床症状。
VOR的功能在于当头部短暂运动时,产生补偿性的共轭眼动,其方向与头动方向相反,从而保持视野清晰,是临床进行前庭功能检查的生理学基础。
由前庭神经节和前庭神经、前庭神经核以及控制眼动的数个核团构成。前庭神经核通过交叉性和非交叉性纤维,经对侧和同侧的内侧纵束,投射到展神经核、滑车神经核和动眼神经核,激活特定的眼外肌和其对应的拮抗肌,引起眼球运动(眼震)。
眼震(nystagmus)是一种不受主观意志控制的眼球节律性运动,前庭性眼震的特征是有交替出现的慢相(slow phase)和快相(quick phase)成分。慢相指眼球向某一方向做相对缓慢运动,由前庭刺激所致;快相则为眼球的快速回位运动,是中枢自发性矫正运动。眼震的慢相一般朝向前庭兴奋性较低的一侧,而快相则正好相反。
VOR主要感受头部的低速和低加速度运动,因刺激前庭诱发,依赖小脑进行精细调节。
VOR可分半规管-眼反射(canal-ocular reflex)和耳石器-眼反射(otolith-ocular reflex),分别产生aVOR和tVOR。此外,头部静止时,在重力加速度的持续作用下,眼球会有微小的反向转动,即视反转(ocular counterrolling),也是一种由耳石器介导的VOR。
1)半规管-眼反射:
刺激外半规管产生水平向aVOR,刺激前、后半规管,产生垂直向aVOR。
A.水平向aVOR:头部向一侧转动,刺激同侧外半规管,输入信号经前庭神经到达前庭神经核,再交叉到对侧展神经核,激活该处运动神经元和中间神经元,导致该侧眼外直肌收缩(即被刺激的前庭对侧);通过内侧纵束到动眼神经核,激活对侧的眼内直肌(即被刺激的前庭同侧)。某一眼外肌兴奋的同时,前庭神经核还会发出抑制性投射到它的拮抗肌,例如同侧眼外直肌兴奋的同时,则对侧眼内直肌被抑制。此外,在头部向一侧转动,则对侧外半规管被抑制,如头向左转时,则右侧外半规管被抑制,其神经元放电频率降低,有助于拮抗肌松弛。
B.垂直向aVOR:假想沿一侧垂直向的直肌和对侧斜肌连成一轴线,则该侧前半规管的走向与之一致。刺激前半规管,通过前庭神经核和2个动眼神经亚核,使同侧上直肌、对侧下斜肌兴奋收缩。刺激后半规管,致对侧下直肌、同侧上斜肌兴奋。所以,刺激垂直向的半规管,所产生的眼震是既有垂直向改变,又呈旋转性。例如,刺激右后半规管,其眼震快相向上,同时眼球向受试者的右侧旋转。因为滑车神经核、动眼神经上直肌亚核控制对侧眼外肌,所以从前庭神经核到数个动眼核团的垂直向神经投射中,兴奋性投射的会越过中线交叉到对侧,而抑制性投射不交叉。
2)耳石器-眼反射:
是指刺激椭圆囊或球囊,信号传入到前庭外侧核,再经脑干投射到对侧展神经核,产生补偿性眼动。共有两种类型:①tVOR,对头部线性加速度的反应;②静态旋转性VOR(static torsional VOR),对头部受持续重力加速度作用时,产生反向的抗旋转性眼动,帮助眼球对抗外力。
主要作用是①延长来自壶腹嵴的前庭输入信号,使aVOR的持续时间延长;②帮助中枢区别直线加速度和重力加速度运动。速度存储机制功能异常,患者出现周期变化性眼震(periodic alternating nystagmus)。
刺激前庭可产生脊髓反射,通过调节颈部、躯干及四肢的肌张力和运动来稳定头部和身体。前庭脊髓外侧束主要支配同侧颈部、躯体及上下肢伸肌收缩。前庭脊髓内侧束主要支配颈髓,进而支配头位。前庭脊髓反射受小脑和高级神经中枢的支配。
对于持续存在或反复给予的刺激,前庭系统出现反应性降低或消失的现象,称疲劳(fatigue)。疲劳程度随刺激强度加大而增强。
前庭习服(habituation)指前庭系统由于受到一系列相同的刺激所表现为反应性逐渐降低或衰减的现象。前庭适应(adaption)是指VOR系统对任何改变了的刺激,进行相应调整,以获得最佳的VOR反应。
代偿(compensation)是指单侧迷路功能急性丧失所引起的眩晕和一些运动症状,可在数日至数周内消失。
冲动复制(pattern-copy)是指当机体受到复杂而有节律的综合刺激时,中枢神经系统即可将这种传入的前庭冲动作为模型加以复制,以便加以对抗和控制。在刺激消失后,这种前庭冲动的复制尚可保留数小时至数日,以致外来刺激虽已消失,机体还存在着与受刺激时相似的反应。
(刁明芳 李晓璐)