
下载掌阅APP,畅读海量书库
立即打开



先天性巨结肠症(HD)基本的病理生理特征是远端狭窄段肠管缺乏神经节细胞,从而丧失蠕动功能,导致功能性梗阻。尽管进行了广泛的研究,但HD的病理生理未完全清楚。目前对于肠管痉挛的发生机制或无神经节细胞肠段的强烈收缩尚无明确的解释。
在内脏器官中,消化道是独特的器官。消化道接触各种各样外源食糜的物理和化学刺激,形成功能丰富且能协调运动的肌性器官,从而确保在消化、吸收、排泄过程中,肠内容物有效地混合和推进。胃肠道的正常运动依赖于神经和肌肉的相互作用。
肠壁包含两层平滑肌,即纵肌层和环肌层。纵肌层位于外侧,由沿肠道长轴排列的薄层细胞形成。环肌层位于纵肌内侧,由垂直肠道长轴排列的厚层细胞形成。两层肌肉之间存在丰富的节细胞神经丛。环肌层的内侧是黏膜下层。黏膜下层含有结缔组织、腺体、小血管、次级神经丛、黏膜下丛。菲薄的黏膜下肌层将黏膜与黏膜下层分开。神经丛的神经细胞发出感觉纤维稠密地分布于黏膜。大量的与肠功能相关的内分泌细胞沿肠黏膜排列。
平滑肌细胞呈细长形且有大细胞核,细胞核位于细胞中央。细胞通过缝隙连接,形成一个强大的功能单位。细胞间电刺激信号通过缝隙连接进行传播。肌肉的活动水平取决于肌细胞本身的活动性和神经功能的发挥。膜电位的慢电波周期性地变化控制着肌肉节律性收缩。激发这些慢电波的起搏细胞是Cajal间质细胞(ICC)。
ICC是一种外形呈纺锤形,或呈多突起的间叶细胞。ICC广泛分布于从食管至肛门的胃肠道黏膜下、肌肉内、环肌和纵肌间,形成一个网状结构。ICC表面可表达一种叫C-Kit的跨膜酪氨酸激酶受体,因此,通过C-Kit免疫组织化学(简称“免疫组化”)染色可以进行ICC的定位。ICC作为肠壁的起搏细胞,发出的慢电波在平滑肌细胞间传播。最近研究表明,ICC也可通过存在于神经末梢和肌细胞内ICC之间的突触样连接来调节肠运动神经递质。但是,ICC的整体作用和肠神经系统对胃肠功能的控制作用机制依然不清楚。
除上述讨论的内源性肌性活动和ICC外,自主神经系统也控制消化道的运动。自主神经系统在非意识状态下控制着内脏功能,可分为脑神经(副交感神经)系统、脊神经(交感和副交感神经)系统和肠神经系统(ENS)3个主要方面。脊神经系统传递外源调控信号。ENS是内在神经系统,不但调节肠道的运动功能,而且调节肠道的排泄、血流、免疫和内分泌功能。
内脏的外源神经包括含支配胃肠道上部的迷走神经和内脏神经,以及支配远段肠道的盆神经。迷走神经内的副交感神经支配胃,大多数神经纤维是细胞体位于神经节内的感觉纤维。这些纤维会将胃和其他外周器官的信息传递给中枢神经系统。内脏神经是交感神经,盆神经包含交感和副交感纤维。脊神经的感觉纤维分布于胃肠道和中枢神经系统,细胞体位于脊神经的背根神经节内。
肠神经系统(ENS)存在于胃肠道壁内,由神经元和支持细胞组成。ENS不但可以通过交感和副交感神经影响胃肠运动,而且可以独立发挥作用。ENS含有约10亿个神经元,是最大的自主神经系统,也是唯一与脊髓神经元数量相当的神经系统。在神经节内聚集着神经元胞体。ENS组成两个神经丛,即肠肌间神经丛(Auerbach丛)和黏膜下神经丛(Henle丛)。肠肌间神经丛位于自食管到肛门的全消化道的纵肌层和环肌层之间。黏膜下神经丛又分为独立的两个神经丛。内黏膜下神经丛(Meissner丛)位于黏膜肌层下,外黏膜下神经丛与环肌层相邻。黏膜下神经丛在食管和胃缺如,在肠道明显。这种分布与神经丛的功能有关,肠肌间神经丛主要调节胃肠道的运动功能,而黏膜下神经丛主要与血流、分泌和吸收有关。肌间神经节与黏膜下神经节之间及不同肠段之间的神经元密度不同。典型的肌间神经节明显比黏膜下神经节大。虽然肠神经系统的神经元同样聚集成神经节,但形态上与脑神经形成的神经核不同。相反,每个肠神经节内含不同种类的神经元,而相邻的神经节内尽管神经元数目不同,但可含有同样类型的神经元。
ENS神经元的分类:可根据形态、神经化学或功能进行分类。区分这些特性需要不同的方法,包括光学和电子显微镜、免疫组化、电生理分析、细胞内染色和神经元投射逆行示踪技术等。目前发现共有17种不同类型的神经元,但只有14种功能重要的神经元被确认。
根据形态,神经元可分为Dogiel Ⅰ~Ⅶ型神经元和巨大神经元,大多数神经元为Dogiel Ⅰ~Ⅶ型。Dogiel Ⅰ型神经元细胞体扁平,有许多短、薄片状树突和一根长轴突,是肠道运动神经元。Dogiel Ⅱ型神经元细胞体相对光滑,细胞体上有各种各样的长、短突起。长突起通过节间纤维延伸,经过数层神经节;短突起仅伸向神经节附近。Dogiel Ⅲ型神经细胞除有更多、相对较短的突起外,与Dogiel Ⅱ型细胞相似。
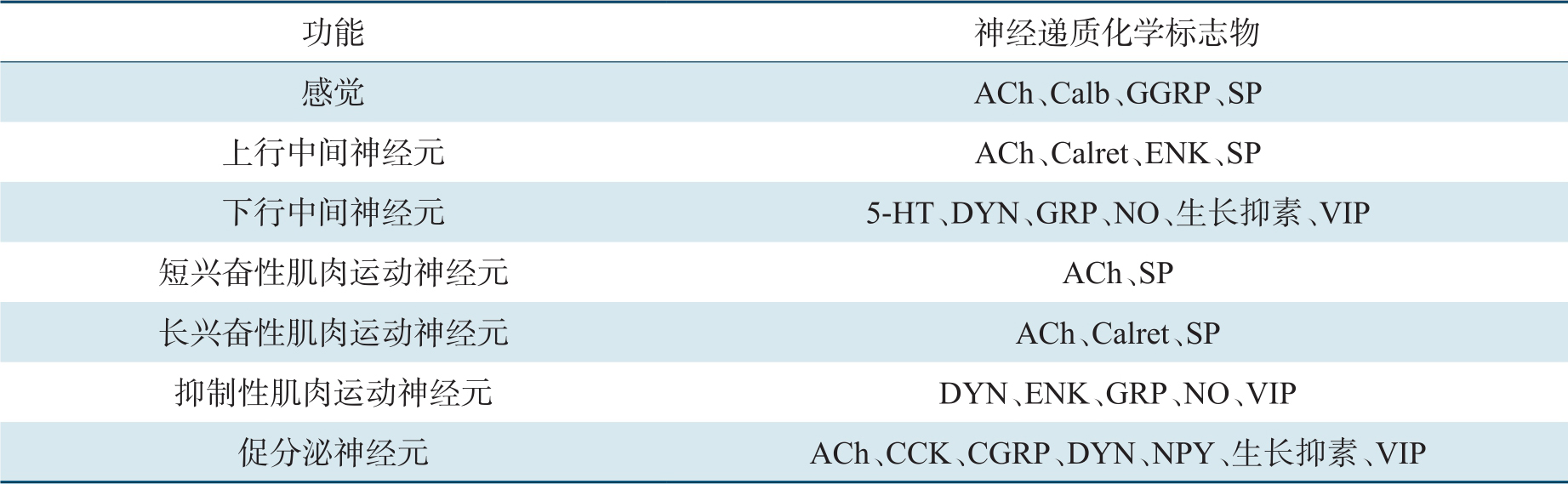
神经元通常分泌不同的神经递质,即化学标志物。神经递质的种类取决于神经元的类型和不同的肠段。ENS中化学递质介导的突触传递机制与其他部位相同,与中枢神经系统的复杂性相类似。ENS已确定释放30多种神经递质,依据功能定位见表2-4。肠神经递质可以是小分子(如去甲肾上腺素、5-HT)、大分子(如多肽),或是气体分子如NO和一氧化碳。
依据功能,神经元分为感觉神经元、中间神经元和运动神经元。
感觉神经元:包括外源性网状传入神经元(细胞体位于肠壁外的迷走神经和脊神经的传入神经)和内源性初级传入神经元(IPAN;细胞体位于壁内)。它们相互交流信息并与内分泌、免疫细胞的功能相互协同。IPAN是ENS控制肠道消化的基础,大脑维持能量、体液的动态平衡及疼痛、不适的感觉亦需要外源性传入神经的信号。感觉神经元包括机械、化学、温度感受器。机械感受器被膨胀信号激活并产生强直性肌肉收缩,但如果膨胀刺激持续,则产生蠕动活动效应。
IPAN除了被直接激活外,还存在特殊的传感器如肠内分泌细胞。这些位于黏膜的内分泌细胞“品尝”和感觉肠内容物,并于基底侧释放调节物,激活固有层中感觉神经末梢,再以突触连接兴奋或抑制性运动神经元传递信号。总之,肠内分泌细胞可特异性地感受肠腔内营养物质,而上皮下的IPAN也可感受肠腔内通过跨上皮自由弥散的化学物质。内分泌细胞在肠道的不同部位分布不同,其中十二指肠最多。主要递质包括胆囊收缩素(CCK)、肠促胰液素、生长激素释放抑制因子、5-HT、促肾上腺素皮质激素释放因子。分泌5-HT的细胞存在于全肠道,是最大的内分泌细胞群体。
中间神经元:通常是Dogiel Ⅱ型神经元,分为一类上行中间神经元和三类下行中间神经元,但大多数是下行神经元。上行神经元为胆碱能神经元,下行神经元分泌复合型递质,包括乙酰胆碱、NO、血管肠多肽、5-HT、生长激素释放抑制因子(表2-4)。
表2-4 肠神经系统神经元的分类
注:5-HT,5-羟色胺;ACh,乙酰胆碱;Calb,钙结合蛋白;Calret,钙视网膜蛋白;CCK,胆囊收缩素;CGRP,降钙素源多肽;DYN,强啡肽;ENK,脑啡肽;GRP,胃泌素释放多肽;NO,一氧化氮;NPY,神经元多肽;SP,P物质;VIP,肠血管多肽。
运动神经元:通常是Dogiel Ⅰ型神经元,包括肌肉运动神经元、促分泌运动神经元(它们是或不是血管扩张剂)和支配肠道内分泌细胞神经元3种类型。肌肉运动神经元支配消化道的纵肌、环肌和黏膜肌层,是兴奋性或抑制性神经元,可释放递质引起肌肉收缩或舒张(表2-4)。兴奋性神经元递质主要是毒蕈碱胆碱能神经元和快速能神经元(P物质和神经激肽A)。抑制性神经元递质主要是NO,也可以是肠血管多肽、ATP、垂体腺苷酸环化酶激动多肽和一氧化碳。
HD典型的病理特征包括狭窄的末端结肠,漏斗状的移行区和扩张肥大的近端结肠。但是,这些特征可以随疾病发展而变化。在新生儿时期,肠道可以表现出正常形态;但随年龄增长,近端结肠逐渐肥大,较正常变厚、变长;结肠袋消失,纵肌层似乎完全包绕结肠。一直以来,HD梗阻症状被认为是继发于远端狭窄段肠管动力异常,但是对于HD远端结肠痉挛发生的确切机制目前依然不明确。
HD远端结肠最特征的表现是在肌间神经丛和黏膜下神经丛中神经节细胞缺失。其中最常见的类型表现为无神经节细胞肠管延伸到直肠和乙状结肠交界区,约占80%。神经节细胞缺乏区肠管是连续无中断的,直达近侧移行区。这一区域的长度可以变化,能延伸至数厘米,以神经节细胞减少症为特征。有关HD其他异常也已经被描述,这些异常改变并丰富了HD的病理生理机制,也可以解释无功能肠道的长度和梗阻程度不同之间的关系。
伴随神经节细胞缺乏症,无神经节细胞肠段的肌间和黏膜下胆碱能神经纤维明显增加。这些纤维表现为粗大的神经干,是外来副交感神经节前纤维。这些副交感神经元连续性释放乙酰胆碱,导致局部胆碱酯酶过量堆积,应用组织化学染色技术能在黏膜固有层、黏膜肌层、环肌中发现这种典型特征。粗大的神经干和增加的胆碱酯酶在无神经节细胞的直肠末端最多,越接近正常肠道越少。近端增加的胆碱能神经纤维分布的肠管范围并不一定与神经节细胞缺乏症程度相对应,通常延伸到更近端。
药理学研究表明,与正常结肠相比,HD无神经节细胞肠段在静息和刺激后释放的乙酰胆碱均明显增加。HD患儿的血清和红细胞中也能发现胆碱酯酶的增多。由于乙酰胆碱是主要的兴奋性神经递质,胆碱能神经纤维增生被认为是无神经节细胞肠段痉挛的原因。但是在神经节细胞缺乏症的动物模型中,应用岩藻胺和升汞后,无神经节细胞肠段没有神经束增生,但肠道仍然狭窄,并出现典型的梗阻症状。由此可见,胆碱能神经纤维的增生并不是肠管出现狭窄痉挛的先决条件。
荧光组化研究用于肾上腺素能神经纤维的定位。研究表明,HD的无神经节肠段肾上腺素能神经纤维数量增加,但分布紊乱。肾上腺素能神经纤维存在于环肌、纵肌及黏膜层,而节细胞正常肠段基本缺如。尽管无神经节细胞结肠段肾上腺素能神经纤维增加,但对肾上腺素的敏感性无明显增加。无神经节细胞结肠段去甲肾上腺素组织浓度是正常肠管2~3倍,调节肾上腺素生物合成的酪氨酸羟化酶也相应地增加。由于肾上腺素的神经生理功能是松弛肠管,所以无神经节肠段肾上腺素能神经纤维的过度活动不是导致肠管痉挛的原因。
NO被认为是参与肠道平滑肌松弛最重要的神经递质之一,它在一氧化氮合成酶(NOS)的作用下,以L-精氨酸和分子氧为底物,合成L-瓜氨酸和NO。NO与胞质中的鸟氨酸环化酶结合,产生3′5′一磷酸环鸟苷酸(cGMP),引起平滑肌松弛。研究表明,NOS与烟碱腺嘌呤二核苷酸磷酸黄递酶(NADPH)的减少共同存在,二者功能相同。
应用NOS免疫组化或NADPH黄递酶组织化学染色,许多研究者对HD患儿无神经节细胞肠段和有神经节细胞肠段的NOS分布进行了检测。在正常人和HD有神经节细胞肠段,黏膜下和肌间神经丛有显著的NADPH黄递酶染色,在环肌、纵肌及黏膜肌层有大量的NADPH阳性神经纤维。而在HD无神经节细胞肠段,肌层及黏膜肌层中NADPH黄递酶阳性神经纤维明显减少或缺如。明显粗大的神经干轻微染色。Kusfuka和Puri检测了7例HD患儿无神经节细胞肠段NOS mRNA,结果显示,NOS mRNA仅为有神经节细胞肠段水平的1/100~1/50。以上结果表明,HD的无神经节细胞肠段中NO合成受损,这种缺陷阻止了平滑肌的松弛,从而导致HD患儿的肠管缺乏蠕动能力。
Bealer等的研究比较了外源性的NO,即S硝基-NO-乙酰-青霉胺(SNAP)对无神经节细胞肠段的等张力平滑肌条的作用效果,结果发现静息张力减少了70%,提示含NOS的神经分布减少可能与HD的发病机制有关。
既往研究表明,包括HD在内的人肠动力性疾病中存在ICC异常。Wanderwinden等首次用C-Kit免疫组化研究HD,发现在HD的无神经节细胞肠段,ICC稀少且分布紊乱,而有神经节细胞肠段的ICC分布与对照组相似。Yamataka等发现,HD肌层C-Kit阳性细胞稀少。在无神经节肠段两层肌肉间的粗大神经干周围有中等量的细胞聚集。Horisawa等发现,C-Kit免疫组化阳性细胞在无神经节和有神经节肠段无差异。Rolle等对全组织包埋切片和冰冻切片进行C-Kit免疫组化染色,发现HD患儿全部切除的肠管中ICC分布的变化不只在无神经节细胞肠段,而且用抗43-连接蛋白抗体定位连接ICC的缝隙连接,发现这种缝隙连接在HD的无神经节细胞肠段缺如,在移行段则大量减少。Rolle等提出HD拖出手术后肠道持续的运动障碍可能与ICC的功能受损和分布改变有关。
应用内分泌细胞免疫组化标记嗜铬素A和突触素,Soeda等证明,HD患者无神经节细胞肠段的内分泌细胞数量比正常肠段明显增加,黏膜中增加的内分泌细胞可以明显影响肠壁的持续收缩,这种收缩主要由5-HT介导。
由于平滑肌是肠运动的最终效应器官,因此,在HD中平滑肌也可能存在异常。平滑肌细胞的细胞骨架由很多蛋白组成,这些蛋白的主要功能是作为结构框架,围绕和支持平滑肌细胞内由肌动蛋白和肌球蛋白丝构成的收缩装置。
Nemeth等应用免疫组化研究平滑肌内的细胞骨架分布,发现在无神经节细胞肠段平滑肌内抗肌萎缩蛋白、纽蛋白和结蛋白免疫反应性缺如或减弱,而在正常肠道或HD患儿的有神经节细胞肠管免疫反应性中等或偏强。神经细胞黏附分子(NCAM)是一种细胞与细胞黏附相关的表面糖蛋白,在神经肌肉系统的发育和功能维持中发挥重要作用。NCAM表达于正常神经支配的婴儿肠管,肠平滑肌某些部分表达密度较低。而在无神经节细胞肠段,平滑肌中NCAM表达出现相反的结果。Kobayashi等发现,与有神经节细胞肠段相比,无神经节细胞肠段黏膜肌层中NCAM表达缺失。而Romanska等发现,在肌层特别是在黏膜肌层NCAM表达增加。以上两个研究均提示在无神经节细胞肠段肥大的神经干NCAM高表达。
目前已经认为细胞外基质(EM)异常与HD的发病有关,EM异常也可能影响到HD的病理生理。应用致死斑点鼠建立远端无神经节细胞症的动物模型,发现平滑肌层中EM成分包括层粘连蛋白、Ⅳ型胶原蛋白、糖胺聚糖和蛋白多糖分布异常。Parikh等证明,无神经节细胞肠段中层粘连蛋白的浓度是有神经节细胞肠段的2倍,是同龄对照组的3倍以上。而且,通过免疫组化方法,发现层粘连蛋白、Ⅳ型胶原蛋白在无神经节细胞肠段固有肌层分布不均衡,且环肌比纵肌表达更明显。同一作者也报道细胞外基质如腱糖蛋白和纤维粘连蛋白在HD患儿的无神经节细胞结肠段表达更明显。
许多研究表明,部分HD患儿手术后很长时间存在持续的肠道运动功能障碍,提示肠道形态和功能异常不仅局限在无神经节细胞肠管。肠神经元发育不良(IND)是ENS的一种畸形,其特征是黏膜下神经丛存在巨大神经节、黏膜固有层存在异位神经节细胞、固有层和黏膜下血管周围乙酰胆碱酯酶活性增强。
1977年,Puri等报道了首例紧靠无神经节细胞肠段近端的IND患儿。此后,许多类似病例相继被报道。一些研究者报道,25%~35%的HD患儿合并IND,并强调这可能是HD拖出术后持续肠道症状的原因。最近,Sandgren等深入研究了致死斑点鼠HD模型近端有神经节细胞肠管,这种动物模型为自然突变的直肠乙状结肠HD模型。他们发现回肠和无神经节细胞肠段近端的结肠黏膜下神经丛中的神经元数量增加,类似于人类的IND。他们认为这些发现可能解释HD术后持续性的肠道运动功能障碍。Sandgren等也表明,在近端有神经节细胞肠段的NO增多且肠血管多肽表达上调,而P物质的表达下调。
20世纪40年代,Swenson等描绘了HD病例肠管标本的蠕动过程图,发现扩张的近端结肠收缩运动并不进入远端的狭窄段,提示远端结肠存在生理缺陷。该发现促进了对此类患儿新的治疗方法即直肠乙状结肠切除术的产生。Kobuta等对HD切除标本的不同肠段的电生理和药理特性进行了多年研究,发现在扩张的有神经节细胞肠段,大多细胞在单一刺激下足够引起一个细胞膜快速超极化继而出现尖峰电压,在移行段超极化的幅度减弱,必须反复刺激才可引起反应,而在狭窄的无神经节细胞肠段反复刺激也只能引起约20%的细胞去极化,只有增加脉冲数才能产生尖峰电位。该研究也已证明阿托品可消除HD全部肠段去极化反应,膜超极化对胆碱能和肾上腺素能神经纤维阻滞剂敏感,并可完全被河豚毒素消除。
以上结果表明肠管存在非肾上腺素能非胆碱能神经抑制性神经支配。通过研究非肾上腺能非胆碱能神经抑制性接点电位振幅变化发现,无神经节细胞肠段接受两种不同来源的神经支配:①来自有神经节细胞肠段经过移行区的内源性抑制性神经支配;②来自低位末端无神经节细胞肠段的外源性兴奋性神经支配。由于移行区是肠内容物停滞发生的部位,因此,内源性抑制性神经冲动的减弱可能是肠梗阻的原因。
总之,虽然HD的组织学特征是神经节细胞的缺如,但其不可能是肠壁痉挛诱发功能性肠梗阻的唯一原因。在HD无神经节细胞和近端有神经节细胞肠段,组织学发现可以解释病变肠管的长度和梗阻的程度不成正比,以及HD拖出术后持续存在的梗阻症状。
(李帅)