
下载掌阅APP,畅读海量书库
立即打开



1.平滑肌细胞的结构特点
平滑肌属于非随意肌,其舒缩活动受到自主神经的调控。平滑肌的紧张性收缩可对抗重力或外加负荷,保持器官的正常形态,并借助于平滑肌收缩而实现其运动功能。
平滑肌细胞呈细长纺锤形,长20~500μm,直径1~5μm。与横纹肌有以下不同:①平滑肌细胞内的细肌丝数量远多于粗肌丝,其比值为(10~15)∶1(横纹肌为 2∶1);平滑肌细胞内没有肌原纤维和肌节样结构,故不显横纹。②平滑肌细胞没有Z盘,作为细肌丝附着点和传递张力的结构是致密体(dense body)和附着于细胞膜的致密斑(图5-2-1)。③平滑肌内的粗、细肌丝斜向走行,呈对角分布,可使平滑肌细胞收缩时呈螺旋扭曲式缩短。④平滑肌细胞的粗肌丝以相反的方向在不同方位上伸出横桥,可使不同方位的细肌丝相向滑行,使粗肌丝和细肌丝之间的滑行范围延伸到细肌丝全长,平滑肌缩短程度达80%(横纹肌不足30%)。⑤平滑肌细胞没有横管,但肌膜上有相应的小凹(caveola),小凹内的细胞外液含有较多Ca 2+ ,小凹的膜上有钙通道。平滑肌细胞的肌浆网位于与小凹对应的胞质内,这是平滑肌细胞实现兴奋收缩偶联的基础。平滑肌的肌浆网不及横纹肌发达,但其膜上存在两种钙释放通道:对三磷酸肌醇(inositol triphosphate,IP 3 )敏感的 IP 3 受体(IP 3 receptor,IP 3 R)和对 Ca 2+ 敏感的雷诺丁受体(ryanodine receptor,RyR)。另外,平滑肌细胞没有内陷的T管结构,因此平滑肌细胞膜上产生的动作电位无法迅速到达深部,这可能是平滑肌收缩缓慢的因素之一。
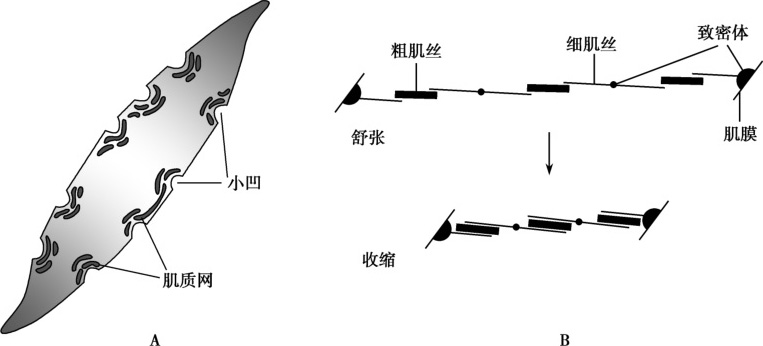
图5-2-1 平滑肌的结构和肌丝滑行示意图
A.平滑肌细胞膜上的小凹及与其对应的肌质网;B.平滑肌细胞内的粗、细肌丝排列和收缩时的肌丝滑行示意图。
2.平滑肌细胞的生物电现象
平滑肌细胞的静息电位在-50~60mV,主要是由于平滑肌细胞膜对Na + 通透性较高所致。平滑肌细胞动作电位的形成主要与钙通道激活有关,由于钙通道的激活和失活都较慢,因此其动作电位的时程较长,一般为10~50毫秒,为骨骼肌的5~10倍。
3.平滑肌细胞的收缩机制
Ca 2+ 是触发平滑肌细胞收缩的关键因子(Wray et al.,2019)。如图 5-2-2 所示,一方面平滑肌细胞可以通过兴奋收缩偶联途径升高胞内Ca 2+ 浓度,即平滑肌细胞在化学信号或牵拉刺激的作用下产生的动作电位可传导至小凹的膜上,进而激活电压门控钙通道,升高胞内Ca 2+ 浓度(主要途径);进入胞内的Ca 2+ 通过与肌浆网上RyR结合引起肌浆网中Ca 2+ 也释放到胞内,只有少量Ca 2+ 是由肌浆网释放的。另一方面,平滑肌细胞还可以在不产生动作电位的情况下接受化学信号而诱发胞内Ca 2+ 浓度升高,即 激动剂收缩偶联(agonist-contraction coupling)途径。如前所述,除了RyR外,平滑肌细胞肌浆网上还存在IP 3 R。体内很多化学信使,包括兴奋性神经递质、激素或药物等,经细胞膜上G蛋白偶联受体-磷脂酶C途径,可使胞内第二信使IP 3 水平升高,进而作用于IP 3 R,导致肌浆网释放Ca 2+ ,是胞质内游离Ca 2+ 浓度升高。在通过肌浆网释放Ca 2+ 活动中,IP 3 R参与的作用更为重要。
与横纹肌收缩机制相似的是,平滑肌收缩也是肌球蛋白和肌动蛋白相互作用下发生的肌丝滑行,由胞质内Ca 2+ 水平增高触发,依赖于ATP供能。但不同的是,Ca 2+ 调控平滑肌收缩的靶点在其粗肌丝上,而不是细肌丝(横纹肌收缩时Ca 2+ 调控靶点位于细肌丝的肌钙蛋白)。平滑肌细胞内不含肌钙蛋白,但有钙调蛋白(calmodulin,CaM),当平滑肌细胞胞质内Ca 2+ 水平升高时,4个Ca 2+ 与胞质内一个CaM结合形成钙-钙调蛋白复合物(Ca 2+ -CaM)。平滑肌细胞的粗肌丝也是由肌球蛋白构成,但其横桥部的ATP酶活性很低,需依赖胞质内的肌球蛋白轻链激酶(myosin light chain kinase,MLCK)来提高其活性。当平滑肌细胞内Ca 2+ 浓度升高时,Ca 2+ 与CaM形成复合物,后者结合并激活胞质内MLCK,活化的MLCK可使肌球蛋白横桥中的调节轻链发生磷酸化,进而引起横桥构象改变及ATP酶活性升高。激活后的横桥与肌动蛋白发生结合、扭动、解离、复位、再结合,进入横桥周期,引起平滑肌收缩。反之,当胞质内Ca 2+ 浓度降低时,MLCK失活,肌球蛋白轻链在肌球蛋白轻链磷酸酶(myosin light chain phosphatase,MLCP)的作用下脱去磷酸,从而使横桥ATP酶活性降低至静息水平并与肌动蛋白解离,平滑肌舒张。
如前所述,静息时平滑肌横桥的ATP酶活性较低,Ca 2+ 与CaM结合并激活胞质中的MLCK后才能提高横桥ATP酶活性,作用于粗肌丝上的收缩调控靶点,因此平滑肌的收缩过程较为缓慢。此外,平滑肌横桥上的ADP释放较慢,故横桥周期中横桥扭动频率远低于横纹肌。横桥周期长的特点使平滑肌收缩时ATP的消耗很少,这对于平滑肌维持长时间的紧张性收缩来十分有利。
根据肌细胞之间的相互关系和功能活动特征,可将平滑肌分为单个单位平滑肌和多个单位平滑肌(图5-2-3)。前者又称为内脏平滑肌,子宫平滑肌属于此类。
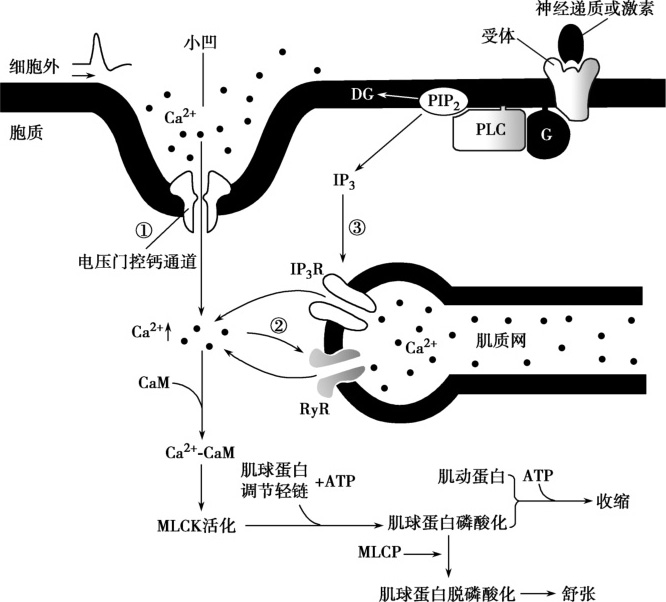
图5-2-2 平滑肌细胞胞质内Ca 2+ 浓度升高与收缩蛋白激活途径
兴奋收缩偶联由细胞膜上电压门控钙通道引起,激动剂收缩偶联经细胞膜上G蛋白偶联受体引起。①兴奋收缩偶联由细胞膜上电压门控钙通道引起;②钙诱导肌浆网钙释放引起的收缩;③激动剂-收缩偶联经细胞膜上G蛋白偶联受体引起。
G.G蛋白;PLC.磷脂酶C;PIP 2 .二磷酸磷脂酰肌醇;DG.二酰甘油;IP 3 .三磷酸肌醇;IP 3 R.三磷酸肌醇受体;RyR.雷诺丁受体;CaM.钙调蛋白;Ca 2+ -CaM.钙-钙调蛋白复合物;MLCK.肌球蛋白轻链激酶;MLCP.肌球蛋白轻链磷酸酶。
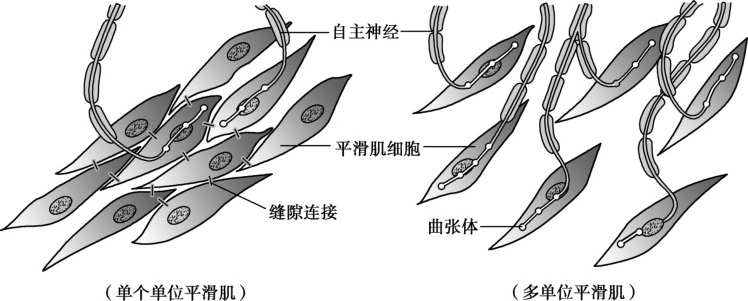
图5-2-3 单个单位平滑肌和多个单位平滑肌示意图
子宫平滑肌细胞间存在大量缝隙连接(gap junction),尤其是妊娠晚期,这是足月或早产分娩发动机子宫同步收缩的先决条件。缝隙连接(图5-2-4)是广泛存在于除血细胞和骨骼肌细胞外的一种组织细胞间连接方式,呈斑点状分散局部连接。电镜下可见细胞间隙仅2~3nm,冷冻蚀刻电镜可观察到相邻细胞膜上有许多规律排布的柱状颗粒,称为连接小体(connexon),每个连接小体由6个杆状的连接蛋白(connexin,CX)构成,中央围成直径约2nm的亲水小管,称中央小管(central canaliculum)。相邻细胞膜两侧的连接小体彼此对接,两侧中央小管连通而形成细胞间直接交通的孔道。这种连接方式构成了相邻细胞间直接进行物质交换的通道,离子和小分子物质(如氨基酸、葡萄糖、激素、信号分子cAMP等)可通过中央小管相互交换,传递化学信息,协调细胞代谢;另一方面,缝隙连接处的电阻低,带电荷的离子可自由通过,有利于动作电位迅速而精确地从一个子宫平滑肌细胞迅速传递给其他平滑肌细胞,实现子宫肌功能和代谢的同步性。这样,子宫的全部平滑肌细胞就能作为一个整体,在化学因素或者机械性因素的调节下产生兴奋性的改变进而进行同步舒缩活动,形成所谓的功能合胞体样活动。与心肌类似,缝隙连接孔道数量越多,其传导性越好。
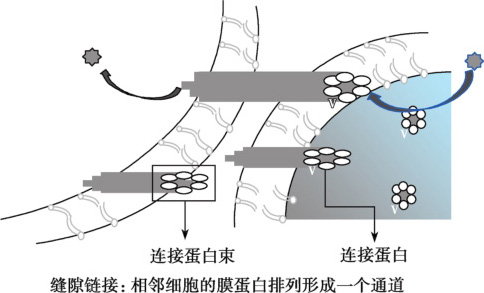
图5-2-4 缝隙连接示意图
缝隙连接是一种动态性结构,其形成机制复杂,膜电位、pH、离子浓度、激素、机械牵拉和细胞黏附分子等可以调节连接蛋白的转录和翻译过程,并可调节其向细胞膜运输、组装形成功能性连接小体的全过程。另一方面,缝隙连接也可以通过变小甚至关闭来保护细胞或是整个组织的损伤。
研究表明,人类子宫肌连接蛋白43(CX 43 )mRNA水平和蛋白水平在分娩启动时显著升高,与机械牵拉和激素水平改变有关。缝隙连接的半衰期为1~2小时,因此在分娩过程中,连接小体处于持续合成状态。CX 43 胞内段的丝氨酸和酪氨酸位点被磷酸化后可易化缝隙连接的通透性,利于子宫肌的同步收缩。而当子宫肌细胞内Ca 2+ 浓度和cAMP水平升高时,缝隙连接的数量就会减少。
与神经肌肉接头间特化的终板样结构不同的是,支配内脏平滑肌的神经纤维在进入平滑肌组织后多次分支,分支上形成许多念珠样的曲张体(varicosity),内含大量囊泡,是递质储存的场所和释放的来源(图5-2-5)。曲张体和内脏平滑肌细胞之间的距离较大,可达100nm,肌细胞膜上的受体散在分布在整个细胞膜表面。因此,神经和内脏平滑肌之间的兴奋传递耗时较长。值得注意的是,外源性神经冲动可以调节内脏平滑肌的兴奋性并影响平滑肌收缩的强度和频率,但是各种体液因子才是调节平滑肌舒缩功能活动的主要因素。如前所述,一些体液因素可以在平滑肌细胞不产生动作电位的情况下,通过影响肌膜上的钙通道或者肌质网上的IP 3 R发挥作用。
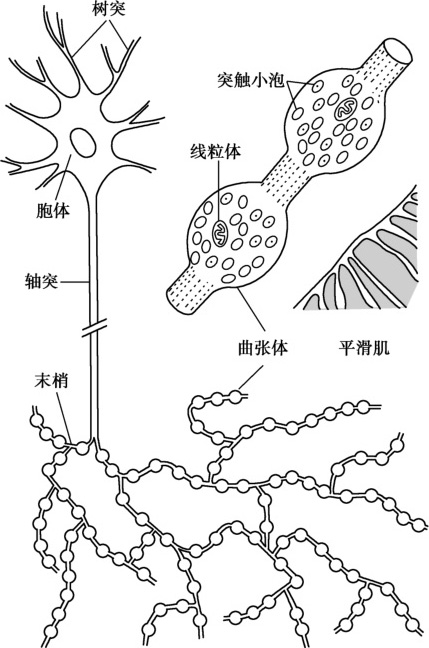
图5-2-5 曲张体和平滑肌结构模式图