
下载掌阅APP,畅读海量书库
立即打开



业已明确,感染是严重烧(创)伤患者的主要死亡原因,其中以全身性感染对患者生命威胁最大。近30年的研究表明,内毒素血症与细菌感染关系密切,它是感染中的重要致病因素之一。实际上,在感染并发症的发病过程中,内毒素常常与细菌协同致病,两者同时并存。尽管感染诱发脓毒症、多器官功能障碍综合征(MODS)的确切机制尚未完全清楚,一般认为细菌内毒素对其发生、发展可能具有促进作用。许多资料揭示,内毒素具有极广泛而又复杂的生物学效应,脓毒症、MODS病理过程中出现的失控炎症反应、免疫功能紊乱、高代谢状态及多脏器功能损害均可由内毒素直接或间接触发。
内毒素是革兰阴性(G - )菌细胞壁的最外层结构,其主要化学成分为脂多糖(lipopolysaccharide,LPS),系脂类、多糖、蛋白质的复合物。脂多糖由三部分组成,外层为O-特异性多糖链,为细菌的特异性抗原;中层为R-核心多糖,其内部核心含有革兰阴性菌LPS所特有的庚糖及2-酮-3-脱氧辛酮糖酸(KDO),为细菌类属共同的抗原;内层为类脂A,主要决定其生物活性。类脂A是LPS的生物活性中心,是一种于氨基和羟基处连接有脂肪酸的二氨基葡萄糖。虽然内毒素的毒性作用和强度随菌种而异,但因各菌属的类脂A结构基本相似,因此不同革兰阴性菌感染时,由内毒素引起的机体反应及临床表现均十分相似。
内毒素的生物学活性多种多样,尤其在体内的作用错综复杂,参与了机体许多病理生理反应过程,主要包括:①刺激单核-巨噬细胞、内皮细胞、粒细胞等合成、释放一系列炎症介质[特别是肿瘤坏死因子-α(TNF-α)、白细胞介素-1(IL-1)]、蛋白酶类物质等,介导机体内多种组织、细胞的损伤;②促进血小板凝集,激活凝血、纤溶系统,从而触发弥散性血管内凝血(disseminated intravascular coagulation, DIC);③对免疫系统的影响:可激活补体、促进B淋巴细胞有丝分裂、诱导干扰素(interferon, IFN),并有抗肿瘤及免疫佐剂作用;④引起机体一系列的病理生理改变:如发热反应、血压降低、代谢改变、局部过敏反应等。目前认为,这些效应主要是通过LPS与细胞表面的受体结合后产生细胞刺激作用,活化细胞内信号转导通路,合成、释放细胞因子及炎症介质,导致体内异常的炎症反应等。
内毒素被发现至今已有一百多年历史,一般认为内毒素只有在菌体破坏(菌体自溶或人工方法使细菌裂解)时才能释放出来。随着对细菌内毒素研究的深入,人们发现了许多革兰阴性菌在其生长过程中亦可以“出疱疹”的方式持续释放内毒素。
关于内毒素的吸收、迁移途径,研究提示胃肠道细菌释放的内毒素可通过被动扩散或主动运输由肠壁吸收入血。Jacob等报道过人体门静脉内毒素血症发生率相当高。肠源性内毒素通过门静脉进入体循环,经肠道淋巴管进入淋巴系统,或由肠黏膜入腹腔。过去普遍认为,门静脉是内源性内毒素的主要吸收途径,然而近年的资料表明,在鼠、狗等动物中,肝系统的支路胸导管是肠道内毒素迁移的重要通道,并且淋巴循环途径的作用及病理生理意义日益受到关注。至于在人体,除门脉系统外的其他迁移途径尚未开展详细研究。
机体对内毒素有清除和解毒作用,使之失活不易造成明显的损害。肝是内毒素灭活、清除的主要部位,通过单核-巨噬细胞系统中库普弗细胞发挥作用。有资料证实,非肠道进入机体的内毒素可在血液中解毒,但其具体机制有待进一步研究。另据报道,中性粒细胞、血小板可能参与内毒素的解毒作用,胆汁在肠道中亦可以灭活内毒素以减少其吸收。正常状态下,因存在上述灭活作用,机体不会发生明显的内毒素血症;但在严重感染或外科应激情况下,体内灭活、清除的功能受损或过量内毒素释放入血,则可能出现内毒素血症。
内毒素血症按其来源不同可分为两类,其一为来自创面或伤道的革兰阴性菌感染或输注含热原污染液体而致外源性内毒素血症;其二是由体内菌库尤其是胃肠道细菌代谢产生并大量释放入血所致的内源性内毒素血症。在严重感染、休克、消化系统疾病、大手术等应激状态下,可能出现如下变化,导致内毒素血症的发生:①全身网状内皮系统功能障碍,免疫功能下降,肠道吸收的内毒素过多超过机体清除能力;②胃肠道黏膜缺血、坏死,屏障功能破坏,通透性增高,大量内毒素释放入血;③肠道来源的内毒素因肝功能障碍由侧支循环直接入体循环;④某些组织、器官革兰阴性菌感染病灶产生的内毒素持续吸收入血。有人曾证实,内毒素血症可发生于肠道缺血的早期,此时还没有细菌的移位(translocation)。许多学者的研究表明,内毒素血症的形成较菌血症为早,可单独存在,造成机体的损害。
烧伤后内毒素血症的发生过程有一定的规律性。有人对3组共39例不同烧伤面积患者在伤后1~5d连续进行血浆内毒素定量测定,结果表明:患者于烧伤后早期即出现内毒素水平升高,伤后3~4d达高峰;循环内毒素水平随着烧伤面积的增加而增加,小于20%总体表面积(TBSA)的患者平均总内毒素量为360内毒素单位(EU),21%~40%TBSA及大于40%TBSA者分别为970 EU、1350 EU。这些资料清楚地说明烧伤后易发生内毒素血症,其含量与烧伤面积相关。我们采用显色基质法鲎试验对35例TBSA大于30%的患者血浆内毒素水平的变化进行了动态观察,结果亦证实,严重烧伤早期血浆内毒素含量即显著升高,以后逐渐下降,2~3周时又出现明显回升。这些患者的烧伤面积为30%~98%,其内毒素血症的发生率相应达36.8%~74.7%,而菌血症阳性率仅3.0%~38.2%(表6 -1),该临床观察结果说明内毒素血症是严重烧伤后较为常见的病理现象。另据第三军医大学烧伤研究所报道,12例严重烧伤并发内毒素血症的患者中死亡组4例患者血浆内毒素水平波动在105~571pg/mL,明显高于存活组(30~240pg/mL),是正常组的16.3倍。根据死亡组患者死前血浆内毒素浓度,推测内毒素血症的预后“线”波动在(325±166)pg/mL,血浆浓度超过此值时,有可能导致死亡。相反血中内毒素水平降低者,全身中毒症状、体征多随之减轻,病情好转,预后较好。总之,尽管在认识上尚有分歧,但多数学者认为内毒素血症是较大面积烧伤后常见的病理过程。
表6-1 不同烧伤面积内毒素血症、菌血症阳性率的比较
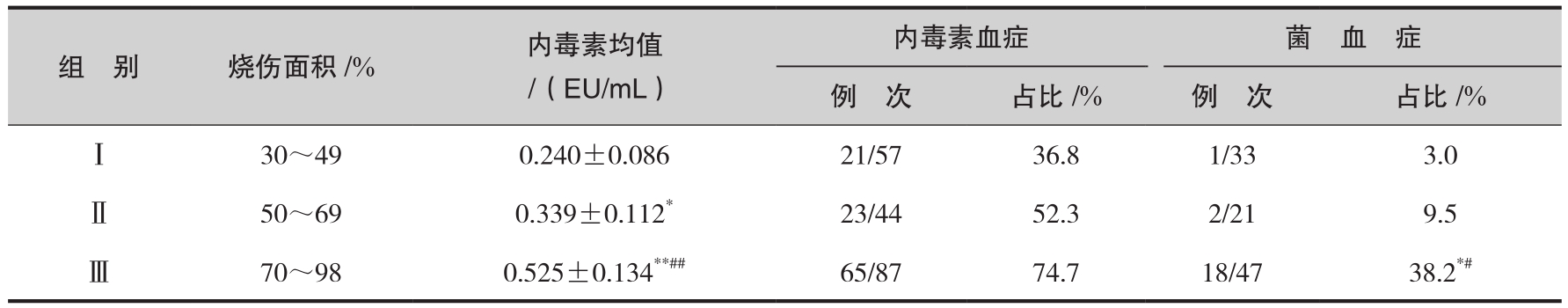
注:与Ⅰ组相比: * p <0.05, ** p <0.01;与Ⅱ组相比: # p <0.05, ## p <0.01。
既往多认为烧伤后内毒素血症来源于烧伤创面或血循环中G - 菌感染后大量释放,近30年来,随着外科领域中肠源性感染研究的深入,人们对烧(创)伤后内毒素血症产生途径有了新的看法。在较大面积烧伤的早期,患者血浆内毒素水平即显著升高,常表现出明显的脓毒症症状,而此时烧伤创面并无大量细菌繁殖,或者血培养无细菌生长。这些现象提示,烧伤早期的内毒素血症主要不是起源于创面,肠道蓄积的内毒素过量侵入血循环或淋巴循环则可能是最重要的来源。临床观察显示,伤后7~12h循环中内毒素含量达峰值,另一高峰则出现在伤后第4天。这种早发的内毒素血症主要来源于肠道,与创面细菌无关,其后内毒素的再度上升则可能与创面脓毒症有联系,因为伤后5d内早期切痂者可显著降低循环内毒素含量。另有资料证实,烧伤面积超过60%的5例患者,伤后1d鲎试验均呈阳性反应,并于烧伤后1~4d内出现MODS,均未发现明确的感染灶。上述诸多临床资料均不同程度地说明烧伤早期可发生肠源性内毒素血症,肠源性内毒素(及细菌移位)可能是导致脓毒症、MODS的重要原因之一。至于烧伤稍后期血浆内毒素水平的再度升高,则与难以控制的创面脓毒症密切相关。
业已证明,大鼠重度出血性休克仅30min,1/3的动物已经出现肠源性内毒素血症,至休克2h其阳性率高达87.5%,同时约半数动物伴有菌血症。我们的系列动物实验显示,致伤前大鼠门静脉血中含有微量内毒素,低水平内毒素可能在激活机体免疫系统并使之处于“预激”状态中起一定作用。烫伤后2h血中内毒素含量迅速升高,伤后8h内毒素水平达峰值,体循环内毒素水平明显低于门脉系统内毒素水平,24h门、体循环内毒素含量基本处于同一水平,说明在烫伤早期肠道内毒素即可通过受损的肠黏膜屏障,由门静脉经过肝而进入全身血循环。生理状态下,肝的库普弗细胞和网状内皮系统具有中和、清除毒素的作用;在烧(创)伤应激状态下,由于肝功能受损而削弱了其灭活、减毒作用,从而使肠道中移位的内毒素得以“溢出”进入体循环或淋巴循环而导致内毒素血症。由于内毒素相对分子质量较小,所以内毒素移位较细菌发生早,达到峰值时间短。进一步研究发现,烫伤后肝、脾、肺组织中内毒素含量均明显高于伤前基础值,提示烧伤后进入体内的内毒素主要分布在肝、脾、肺等组织中。而伤后肝组织内毒素含量明显高于脾、肺组织,表明肝脏可能是烧伤后内毒素最主要的分布场所(表6-2)。据报道,给动物静脉注射放射性标记内毒素后5min,99%以上的内毒素即迅速从循环中清除。免疫组化检查显示,从循环中清除的内毒素迅速分布到肝、肺、肾等组织巨噬细胞内。内毒素及其代谢产物经巨噬细胞代谢转运后可逐渐由胆道排泄,采用酚-水抽提法提取从内毒素血症大鼠胆汁中排出的内毒素,发现它对小鼠仍具有极强致死活性,说明内毒素攻击时,尽管循环中内毒素迅速被单核-巨噬细胞系统所清除,但进入组织中的内毒素仍可保留一定的生物学活性。大量动物实验研究证明,严重烧伤后常伴有不同程度的休克期。由于创伤应激状态,往往破坏肠上皮细胞之间的紧密连接,导致肠黏膜屏障功能削弱,肠黏膜通透性迅速增高,从而使肠道中蓄积的内毒素得以侵入机体内形成内毒素血症,循环中的毒素又反馈性促进肠道中内毒素、细菌持续入血,形成恶性循环。所以,对烧伤休克期的治疗,除液体复苏外,还应注意拮抗早期内毒素血症,以避免或减少对机体的继发性损害。
表6-2 烫伤大鼠组织内毒素含量的动态变化
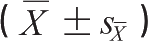 EU/g
EU/g
注:与对照组相比: * p <0.01。
诸多研究表明,严重烧伤、休克、缺血-再灌注损伤等应激状态下发生的脓毒症及多器官损害与肠道细菌-内毒素移位所致肠源性感染密切相关。但是,急性损伤打击后血浆内毒素水平仅呈现一过性增高,脓毒症患者体内内毒素含量一般在pg/mL水平。而离体实验中,pg/mL水平内毒素并不能刺激细胞应答,剂量提高数百倍才能激活细胞。因此,人们推测机体内毒素增敏效应可能参与了严重烧(创)伤后脓毒症、MODS的发病过程。近年来体外观察发现,脂多糖结合蛋白(lipopoly saccharide binding protein, LBP)及脂多糖受体(LBP/CD14)系统是机体识别和调控内毒素作用的关键机制之一,内毒素的许多生物学效应可能是通过其增敏作用来实现的,但我们对其体内的作用了解甚少。我们采用多种动物模型,并结合烧(创)伤临床病例的前瞻性研究,从不同层次观察烧(创)伤后LBP/CD14系统的变化规律及其与多器官损害的关系。通过该系列研究,试图初步明确LBP/CD14系统在烧(创)伤后内毒素移位诱发脓毒症中的作用,并为进一步探讨脓毒症、MODS的分子发病机制和早期干预提供新的理论依据。
肠缺血-再灌注(I/R)损伤是严重烧(创)伤后常见的病理生理过程,是导致脓毒症及MODS的重要诱发因素之一。有资料证实,烧伤、I/R及失血性休克均可显著提高宿主对内毒素的敏感性。据报道,大鼠40%烫伤后肺泡巨噬细胞TNF-α诱生能力显著增强,高达伤前值的11.1倍,表明烧伤可预激与活化机体多种组织巨噬细胞,从而使其对内毒素的敏感性明显提高,但其确切机制尚不清楚。我们采用大鼠急性I/R模型,初步探讨了CD14在介导内毒素所致炎症反应及器官损害中的意义。结果显示,I/R组或内毒素组(LPS组,手术后12h注射LPS 3.0mg/kg)平均动脉压、心脏指数、每搏输出量与假手术组(S组)相近或仅轻度下降,缺血-再灌注合并内毒素组(I/R+LPS组,再灌注后12h注射LPS 1.5mg/kg)则血流动力学异常改变显著加重。除血清尿素氮(blood urea nitrogen, BUN)以外,反映器官功能的指标如谷丙转氨酶(alanine aminotransferase, ALT)、D-乳酸、肺毛细血管通透性等在I/R+LPS组改变最重,其升高幅度均明显大于I/R组或LPS组(表6-3)。体外观察发现,LPS浓度≤10ng/mL时,抗CD14单抗可显著抑制全血TNF-α的产生;但当LPS浓度≥100ng/mL时,其诱导TNF-α的产生受CD14单抗的影响较小。此外,I/R后抗CD14单抗对内毒素诱导TNF-α的抑制效应明显增强,当LPS浓度≤10ng/mL时其抑制率显著高于伤前值或假手术组。
表6-3 大鼠肠缺血-再灌注后内毒素对器官功能指标的影响
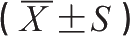
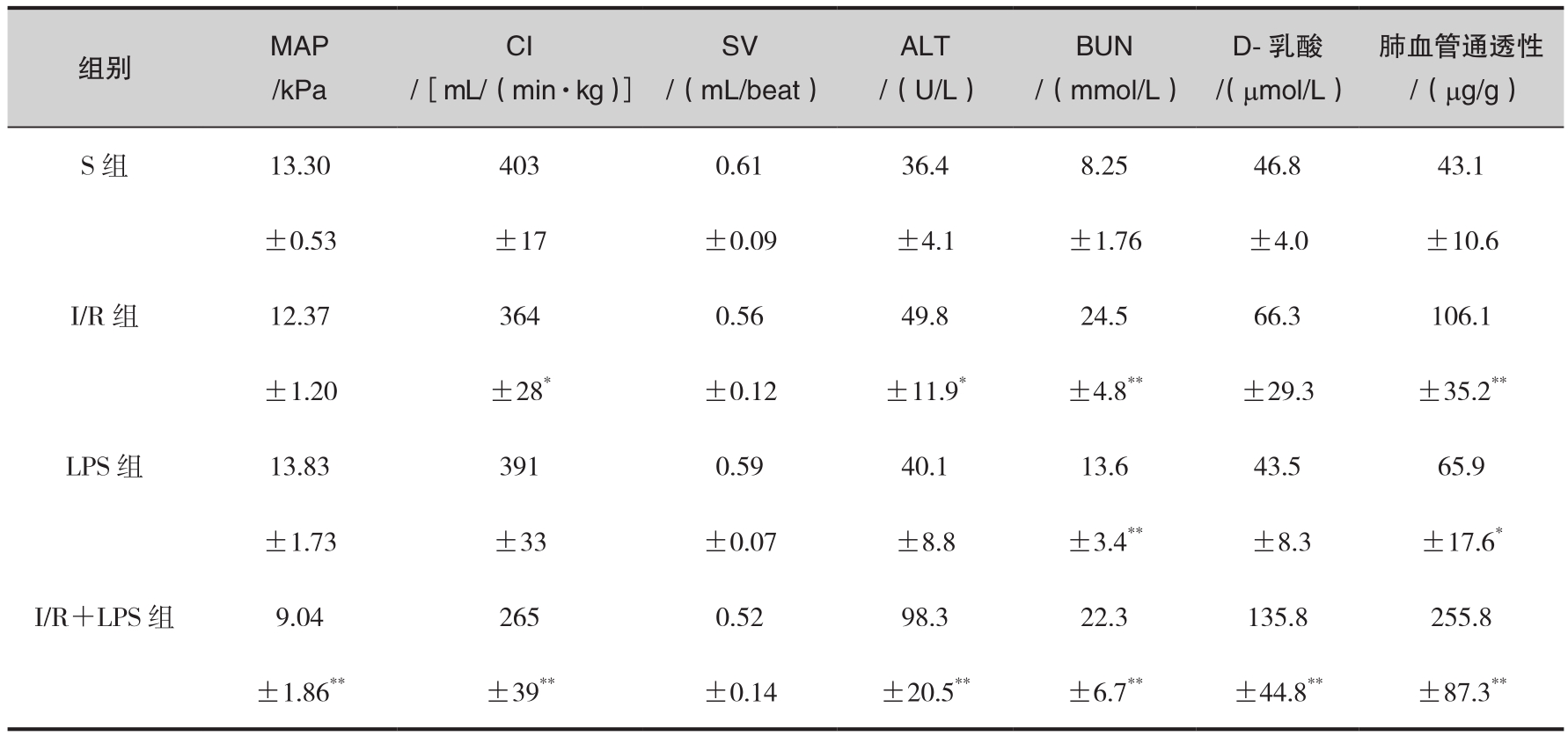
注:与S组比较, * p <0.05, ** p <0.01。
上述结果提示,I/R可显著提高机体对内毒素攻击的敏感性,从而为继发性打击导致脓毒症或MODS起了“预激”作用。关于I/R增敏内毒素作用的机制,推测可能与机体识别和调控内毒素效应的LBP/CD14系统有关。该研究发现,CD14介导细胞应答反应与内毒素刺激剂量密切相关。同时,I/R后内毒素刺激全血产生TNF-α主要依赖于CD14,且体内CD14依赖途径的作用明显增强。因此,I/R、烧伤或失血性休克打击可进一步激活内毒素增敏系统,初步提示该效应与急性损伤后机体CD14表达的上调有关。
动物实验和临床观察揭示,内毒素与严重创伤后一系列感染并发症,如脓毒症、MODS等密切相关,但其详细的分子发病机制仍有待于深入研究,新的防治途径值得进一步探索。在既往研究的基础上,我们采用重度低血容量性休克、烧伤等动物模型探讨LBP在严重创伤后内毒素血症诱发多器官损害中的作用及重组杀菌/通透性增加蛋白21(bactericidal/permeability increasing protein,rBPI 21 )的防治效应。结果显示,重度低血容量性休克180min后血浆内毒素水平明显升高,复苏末达峰值。同样,严重烫伤后2h外周血内毒素水平亦显著升高,8h达峰值。给予rBPI 21 治疗则可完全防止休克所致内毒素血症的发生,烫伤治疗组动物内毒素峰值比对照组降低42.4%。另一方面,大鼠休克、复苏8h后肝、肺、肠组织LBP mRNA表达均显著增强,肾组织表达则呈阴性。rBPI 21 治疗可明显抑制肝、肺组织LBP表达水平,但对肠组织影响不大。同样,烫伤治疗组肺、肠、肾组织LBP mRNA水平明显下调。病理形态学检查显示,休克对照组动物肺、肝、肠、肾等器官病理损害较重,rBPI 21 处理组则相对较轻,病变发生频率亦明显降低。休克对照组及rBPI 21 治疗组48h动物存活率分别为37.5%(6/16)和68.6%(11/16),rBPI 21 组动物预后趋于显著改善。
实验结果表明,严重低血容量性休克、烫伤早期注射rBPI 21 可显著降低循环内毒素水平,从而防止肠源性内毒素血症的发生与发展。与此同时,肺、肝、肾、肠等多种组织LBP mRNA表达不同程度地被抑制,尤其以休克、复苏后肝组织及烫伤打击后肠组织下降最为明显。另一方面,病理形态学检查证实,严重休克后多脏器损害明显减轻,动物病死率比对照组降低31.3%。由此可见,低血容量性休克、烧伤后肠源性内毒素血症在多器官损害的发病过程中发挥了重要作用,及早、有效地阻断急性损伤初期内毒素血症的发生,有助于抑制机体LBP的增敏效应和过度炎症反应,从而降低多个器官对内毒素损害的敏感性,防止脏器功能障碍的进一步发展。
体外观察已经明确,CD14作为LPS的功能受体,可能受内毒素的直接调控或内毒素介导产生的各种介质的间接调节。据报道,LPS可上调人单核细胞CD14 mRNA的表达,同时细胞表面膜CD14(mCD14)分子及培养上清液中可溶性CD14(sCD14)水平均显著增多,并且LPS还可上调猪肺泡巨噬细胞、人中性粒细胞表面mCD14的表达。基于上述体外观察的结果,我们推测烧伤后组织CD14系统表达上调可能与移位的内毒素有关。动物实验结果显示,组织内毒素水平与组织CD14 mRNA表达程度呈显著正相关,提示肠源性内毒素对机体多种组织CD14 mRNA表达具有重要影响。为了进一步明确组织内毒素在刺激CD14 mRNA表达中的作用,我们采用具有中和内毒素效应的rBPI 21 进行治疗,以探讨二者的因果关系。结果显示,rBPI 21 治疗可有效地防止不同组织肠源性内毒素移位的发生,同时,给予rBPI 21 可显著抑制局部组织CD14 mRNA水平。伤后12h,治疗组动物肝、肾、肠组织CD14的基因表达均恢复至伤前范围;肺组织表达水平亦明显减弱,伤后12h其表达量为烫伤对照的43.2%,24h则降至伤前基础值。该结果说明,烧伤早期肠道内毒素发生移位并蓄积于局部组织,它对于进一步上调不同组织CD14基因表达具有重要作用。有资料证实,给小鼠腹腔注射内毒素后,各组织内源性细胞中CD14 mRNA水平明显高于对照组,同时血浆sCD14水平也显著升高,值得注意的是,正常情况下基本不表达CD14的上皮细胞和内皮细胞CD14 mRNA表达亦明显增强。由此可见,内毒素可引起CD14在不同组织中广泛表达,从而为进一步介导器官损伤奠定了基础。这样,机体CD14表达上调可能对增强机体对内毒素的敏感性、诱发全身失控炎症反应及脓毒症具有促进作用。
临床资料显示,14例严重多发性创伤患者血浆LBP含量伤后第1天即明显增多,并呈持续上升趋势。与正常对照组比较[(4.71±1.59)µg/mL],多发性创伤组伤后第3、7天LBP含量显著性升高。进一步分析可见,循环内毒素与LBP水平伤后1、3d均呈显著正相关。该资料表明,急性损伤早期内毒素血症对体内LBP的产生有重要影响。与之相似,在狒狒低血容量-创伤性休克模型中观察到,休克打击后48h、72h血浆LBP均值分别为22.18μg/mL、30.66μg/mL,显著高于伤前基础值(3.88µg/mL),并且循环LBP的合成、释放与肠源性细菌-内毒素移位密切相关。因此,创伤、休克早期抗内毒素治疗可能有助于减轻机体LBP的大量产生,进而部分阻断LBP介导的内毒素增敏效应,这对于防止脓毒症、MODS的发生与发展可能有一定意义。为了探讨血浆LBP水平与患者预后的关系,对14例多发性创伤患者中存活者与死亡者进行比较,发现死亡组LBP水平伤后第3天显著高于存活者。这些结果证明,创伤早期LBP可能参与了机体脓毒症的病理过程,并介导内毒素对不同器官的广泛损害。动态检测循环LBP水平可能有助于患者预后的判断,关于LBP确切的临床预警价值仍有待于深入研究。
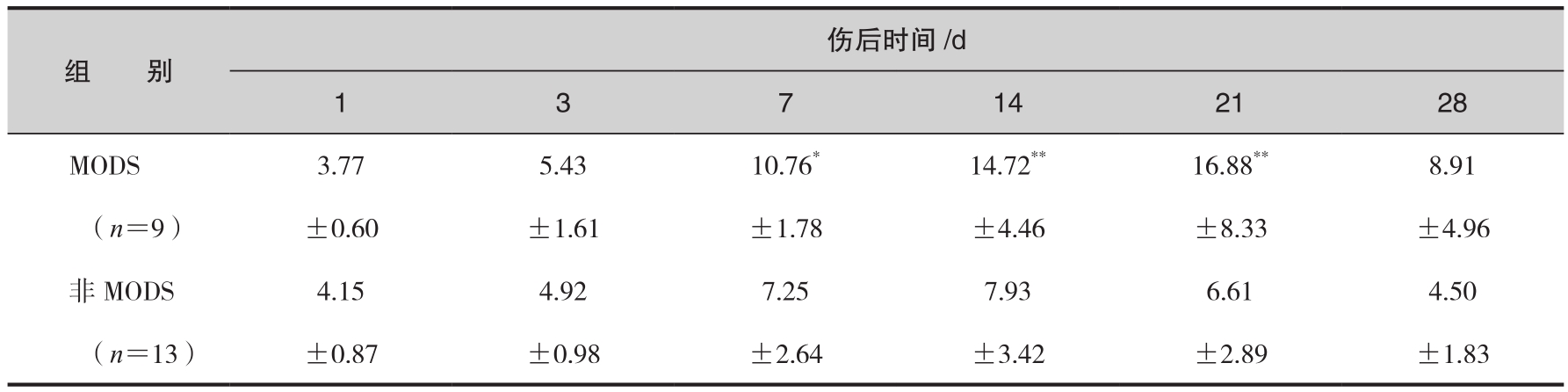
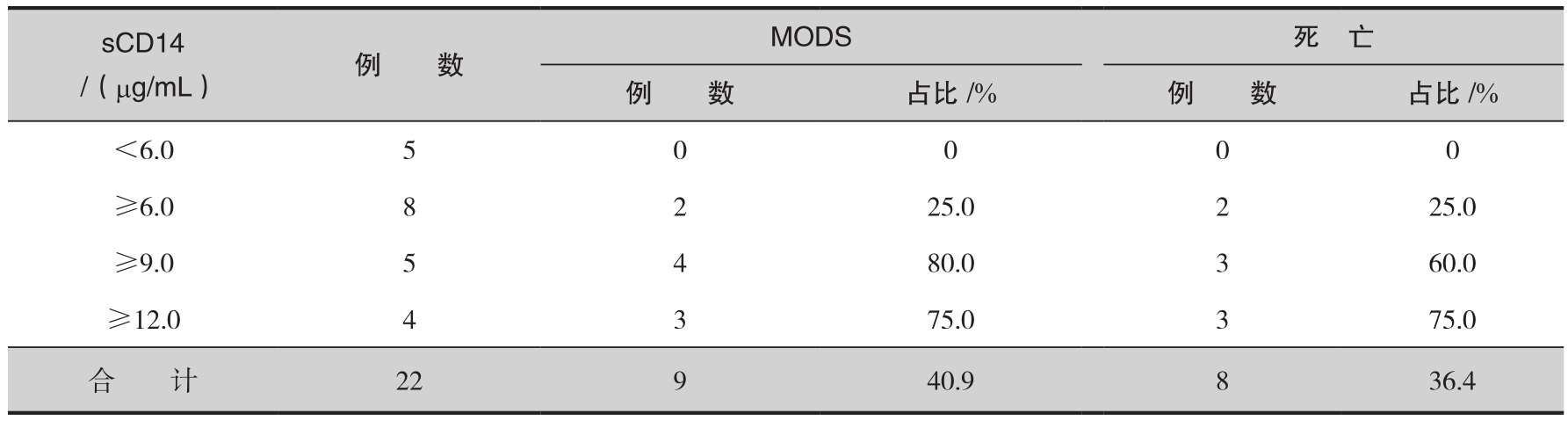
另一组资料观察到,生理及病理情况下血清中均存在一定水平的sCD14。大面积烧伤早期sCD14含量改变不明显,伤后第7天则明显升高,且一直持续至伤后第3周。进一步分析可见,烧伤后第7天、14天和21天MODS组sCD14水平显著高于非MODS组(表6-4),且sCD14升高程度与MODS发生呈平行关系,当其均值超过9.00µg/mL时,9例患者中有7例发生了MODS。这一观察结果提示,sCD14水平与严重烧伤后MODS的发生、发展密切相关,动态监测其改变对MODS的发病过程具有一定的预警意义。值得注意的是,sCD14含量不仅与MODS的发生有关,而且与患者的预后密切相关。该组资料中,随着sCD14均值的不断上升,患者病死率亦逐渐增高。sCD14含量持续于较高水平(≥9.0µg/mL)常为预后不良之兆(表6-5)。由此可见,动态检测sCD14的变化是判断严重烧伤后病情转归较为客观的指标。
表6-4 MODS、非MODS患者血清CD14水平比较
 µg/mL
µg/mL
注:与非MODS组同一时间点比较, * p <0.05, ** p <0.01。
表6-5 sCD14与患者MODS及预后的关系
既往的研究已证实,大面积烧伤可导致内毒素、细菌持续不断侵入体内造成内毒素血症,它对于进一步诱发机体失控的炎症反应及全身多个脏器的损害具有一定作用。一组资料中,我们观察到sCD14水平与烧伤后MODS的发生、发展密切相关,且其改变与循环内毒素呈平行关系。值得重视的是,该组9例MODS患者中,肺部并发症出现时间最早、发生率亦最高,推测与循环sCD14过量弥散、聚集于肺组织有关。最近,有人发现ARDS患者支气管肺泡灌洗液sCD14含量与肺部炎症反应呈高度正相关,局部组织sCD14大量产生、释放可能是导致急性肺损伤的重要原因之一。有鉴于此,烧(创)伤后循环sCD14的持续存在对于促进内毒素血症介导的机体广泛损害可能有一定临床意义。
严重烧伤患者sCD14水平升高的病理生理意义还不完全清楚,其致病机制可能是在LBP的催化下,较低水平LPS与sCD14结合形成复合物,在sCD14的介导下激活CD14阴性细胞,如内皮细胞、上皮细胞等,使其表型发生改变;激活的细胞表达大量黏附分子,释放炎症细胞因子(IL-1、IL-8等),促凝活性增强;同时,在sCD14介导下,LPS直接使内皮细胞通透性增高,造成损伤。这样,sCD14介导的一系列反应促使白细胞浸润和微血栓形成,最终产生广泛的机体损伤效应。关于烧(创)伤后sCD14水平升高的机制正在研究之中,LPS或细胞因子等多种因子均可诱导产生sCD14。我们的临床资料发现,sCD14与血浆内毒素水平呈显著正相关,有人还观察到其与血浆TNF-α水平呈显著正相关。体外实验证实内毒素、TNF-α均可刺激单核细胞产生sCD14,释放到培养上清液中。而脓毒症、MODS时循环内毒素和TNF-α均可升高,因此sCD14的诱生可能与二者的直接或间接作用密切相关。
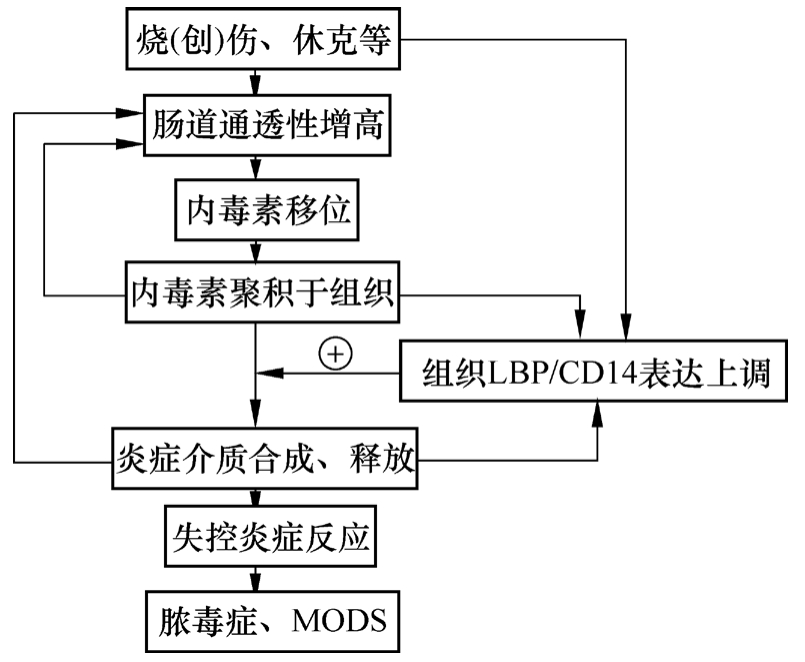
图6-1 内毒素增敏假说
基于上述分析可知,LBP/CD14系统通过介导内毒素刺激组织合成、释放TNF-α等炎症介质,在烧(创)伤后脓毒症、MODS的发病机制中发挥重要作用。而组织LBP/CD14 mRNA表达增加可能是严重创伤增敏内源性内毒素作用,从而发挥其生物学效应的主要分子基础。由此,我们设想烧(创)伤、休克后脓毒症、MODS可能是循以下途径发生、发展的(图6-1):创伤、烧伤、休克等应激状态(首次打击)一方面引起机体LBP/CD14系统表达上调,使机体对内毒素的敏感性增高(致敏阶段);另一方面,由于免疫抑制、肠道通透性增高以及菌群紊乱等因素,促进肠源性内毒素移位,并聚积于局部组织(二次打击)。组织内毒素通过表达上调的LBP/CD14系统激活多种炎症细胞,释放炎症细胞因子(TNF-α等)。同时,内毒素及炎症介质一方面持续上调组织LBP/CD14表达,另一方面则进一步加重肠黏膜损伤,促进内毒素移位。从而在内毒素- LBP/CD14 -炎症介质之间形成一正反馈环,致使炎症反应不断放大、加重,最终导致全身失控性炎症反应和MODS。
内毒素血症与细菌感染关系密切,它是烧(创)伤感染中的重要致病因素之一。实际上,在脓毒症发生过程中,内毒素常常与细菌协同致病,两者同时并存;但有时血液内毒素检测呈阳性反应,而血液细菌培养不一定阳性,提示内毒素血症也可单独存在。有资料表明,严重烧(创)伤、手术应激后肠源性内毒素血症的出现时间往往要早于菌血症的发生,细菌内毒素对肠源性感染具有促进作用。有学者采用无特殊病原菌(specif i c - pathogen free, SPF)小鼠观察了内毒素对蛋白营养不良动物肠道细菌移位的影响。实验发现,大肠埃希菌内毒素血症能显著促进营养不良小鼠合并烧伤后肠道细菌的进一步播散,形成全身性感染,并增加烧伤后动物病死率,而单纯营养不良并不能造成细菌移位的发生。目前认为,应激状况下循环内毒素可促进肠源性细菌移位的发生、发展,其机制可能主要是通过破坏肠黏膜机械屏障及提高肠黏膜的通透性所致。应用电镜组化及冷冻蚀刻技术观察到,内毒素能导致连续上皮细胞之间的紧密连接组织结构发生明显改变;使细胞间“通道”异常开放,肠黏膜对肠腔内细菌、内毒素的通透作用增强;内毒素血症不仅对肠道细菌移位具有促进作用,更值得注意的是,它与严重创伤后全身播散性感染的发生亦紧密相关。根据临床观察报道,内毒素血症与革兰阴性菌脓毒症的相关率达70%以上,当血浆内毒素含量越来越高时,应警惕播散性细菌感染的发生。因此,早期诊断并及时阻断内毒素血症的发生、发展,对于阻止细菌感染的进一步加剧,防止脓毒性休克、MODS并发症具有积极作用。
严重烧(创)伤后内毒素血症对宿主的影响是多方面的,它在体内可引起一系列的病理生理变化与临床中毒症状。一些研究揭示,细菌内毒素在创伤后机体免疫抑制中扮演着较重要的“角色”。较大面积烧伤患者细胞免疫的异常表现为外周血淋巴细胞在体外对细胞促有丝分裂原,如植物血凝素(phytohemagglutinin, PHA)、念珠菌抗原等刺激反应降低,同时伴有抑制性T细胞(suppressor T cell, Ts)数目增多、辅助性T细胞(helper T cell, Th)减少。通过体内、体外试验发现,上述细胞免疫功能障碍均与患者血清中存在过量的内毒素有关。相反,应用较低剂量的多黏菌素B治疗后,可部分恢复患者自然杀伤细胞(natural killer, NK)的活性及逆转Th/Ts细胞比例的严重异常,且能消除烧伤血清对IL-2的抑制反应。由此可见,抗内毒素治疗对改善烧伤后免疫功能的障碍有一定价值。
传统的观点认为创伤、休克时首先导致免疫功能低下,然后才发生细菌及其毒素的侵袭,出现脓毒症或脓毒性休克。近年来,肠源性内毒素对机体全身免疫功能的影响逐渐受到重视。我们的观察证明:休克初期即发生内毒素血症及细菌入血,而通过体内重要免疫抑制因子——前列腺素E 2 (prostaglandin E 2 ,PGE 2 )等介导的免疫抑制作用相对要迟缓,Re-LPS抗血清输注后血浆PGE 2 水平较对照组明显降低。因此,肠源性内毒素血症对血浆PGE 2 水平有一定影响,而后者的大量产生、释放可能与机体免疫功能紊乱、感染的易感性增加密切相关。为了探讨内毒素血症与机体全身性细胞免疫的直接关系,我们还观察了烫伤动物脾细胞对促有丝分裂原增殖应答反应、IL-2活性及T细胞亚群比值等指标的变化。结果发现,大鼠40% Ⅲ度烫伤后,脾淋巴细胞对刀豆素A(Con A)或PHA刺激后增殖反应及脾细胞诱生IL-2活性明显抑制。针对肠道内毒素移位,预防性进行选择性消化道脱污染(selective decontamination of the digestive tract,SDD)后,则可显著降低循环内毒素水平,同时脾细胞的增殖应答及诱生IL-2活性较烫伤组明显升高,但对外周血T细胞亚群比值无显著影响。这些观察提示,严重烫伤应激后肠道中内毒素的过量入血对于诱发全身性细胞免疫功能异常有一定作用。关于细菌内毒素介导创伤后免疫抑制的作用机制,可能主要有如下几方面:①刺激单核-巨噬细胞系统产生、释放大量的PGE 2 ,通过PGE 2 对免疫系统的负向调控(down-regulation)引起多方面的抑制效应,如Th细胞活性增强、单核细胞分泌IL-1增加、Th细胞IL-2产生减少等;②直接或间接作用导致单核细胞呈递抗原表达能力降低,并抑制中性粒细胞趋化性受体的表达;③激活体内单核-巨噬细胞系统诱生TNF-α、IL-1等细胞因子,介导或协同产生对机体防御功能及多器官的损害效应;④经替代途径活化补体,生成多种补体裂解产物,如C3b、过敏毒素等。当然,损伤后机体免疫抑制的机制比较复杂,细菌内毒素作为一个较重要的因素可能参与了这一病理过程。
脓毒症、MODS诱发因素十分复杂,其中革兰阴性菌严重感染为主要原因之一。感染诱发脓毒症、MODS的确切机制尚未完全明了,一般认为细菌内毒素对其发生、发展具有促进作用。大量研究揭示,内毒素具有极广泛而又复杂的生物学效应,脓毒症、MODS病理过程中出现的失控炎症反应、免疫功能紊乱、高代谢状态及多脏器功能损害均可由内毒素直接或间接触发。业已明确,细菌内毒素主要是通过激活机体单核-巨噬细胞系统过度释放中间介质而发挥作用,其中TNF-α、IL-1可能是介导内毒素损害效应、诱发脓毒症的关键早期细胞因子。我们在家兔MODS模型中观察到,内毒素血症与MODS的发生、发展密切相关。并发MODS的动物血浆内毒素含量升高幅度大、持续于较高水平,且内毒素水平的改变与多器官功能指标相关显著。给失血性休克家兔输注Re-LPS抗血清后,血浆内毒素水平的升高幅度及其持续时间均显著降低,动物MODS发生率明显低于对照组(11.2%与58.0%, p <0.01)。同样,重度出血性休克早期给予具有抗菌及抗内毒素双重作用的rBPI 21 ,可完全中和循环内毒素,能有效地减轻肝、肺、肾及肠道损害等。此外,预防性进行SDD大鼠,其各段肠腔中游离内毒素含量较对照组下降99.5%以上,门、体循环内毒素水平随之显著降低。防治组肠黏膜损害减轻,严重烫伤后其存活率提高26.7%。另一方面,我们的临床观察证实,大面积烧伤患者内毒素血症发生率为58%,脓毒症组血浆内毒素均值显著高于非脓毒症组,且血浆内毒素水平与烧伤后MODS发生频率呈正相关。持续严重内毒素血症(≥0.50EU/mL)者,多呈现显著的脓毒症状态,最终可并发MODS而死亡(其中3例患者死前血浆内毒素浓度超过1.00EU/mL)。相反,非MODS者伤后早期尽管血浆内毒素暂时性升高,但其变化趋势进行性下降,致伤1周之后仅表现为轻度内毒素血症,患者感染症状多随之减轻,预后较好(表6-6)。这与我们的动物实验结果是一致的。上述结果证明了肠源性内毒素血症与烧伤后脓毒症、MODS发病可能具有密切关系。
表6-6 严重烧伤后MODS、非MODS患者血浆内毒素含量比较
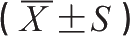 EU/mL
EU/mL
注:与非MODS相比: * p <0.05, ** p <0.01。
创伤后MODS常继发于严重感染,因此有人称之为脓毒综合征。Meakins主张胃肠道是其始动部位,通过细菌和内毒素起作用。近年来,许多学者对于MODS的感染机制开展了深入的研究,提出了细菌内毒素作为重要触发因子的观点,并受到人们的普遍关注。虽然很多人支持内毒素在MODS发生中起着“扳机”作用,但亦有不少研究者提出异议。有人给大鼠腹内注射酵母多糖复制出非细菌感染性多器官损害的动物模型,认为MODS是一种具有广泛炎症特点的全身性自身破坏过程。脓毒并发症是宿主对感染因素的一种全身性反应,其本质及重要性并不在于感染因子(细菌、毒素、病毒等)本身的直接效应,而在于机体的失控炎症反应影响了体内神经-内分泌-免疫网络,最终造成细胞能量代谢紊乱及功能异常。Moore等对20例可能发生MODS的创伤患者进行紧急腹部手术,并行门静脉插管以便连续采集标本,虽然最终有30%的患者并发MODS,但在伤后5d内门静脉血与外周血中均未检测到细菌和内毒素。不少报道也证实,在急性创伤患者中细菌移位至肠系膜淋巴结并不多见,同样,也未发现失血性休克和严重创伤患者血浆内毒素水平显著升高。另一方面,临床上选择性肠道去污术虽然能明显减少感染并发症,但病死率并未降低。因此,肠源性内毒素血症的临床意义有待进一步探讨。
内毒素血症的临床表现大都与革兰阴性菌所致脓毒症相似,对创伤患者内毒素血症的诊断除临床体征外,主要依据血中内毒素测定结果。尽管血液内毒素检测方法尚有待进一步完善,但因其具有快速、灵敏、不受抗菌药物治疗影响等优点而逐渐应用于临床。血浆内毒素含量可以比较稳定和敏感地反映机体革兰阴性菌感染的动态变化过程,内毒素浓度越高,患者全身中毒症状越明显。我们的临床观察表明,并发MODS组患者血液内毒素水平自伤后第3天起明显高于非MODS组,且循环内毒素含量与MODS发生频率呈正相关关系,内毒素水平持续超过0.5EU/mL者,应高度警惕并发MODS的可能。随着循环内毒素含量的不断升高,MODS的发生频率及病死率逐渐增加(表6-7),反之,患者感染症状则多随之减轻,预后较好。另据报道,依据死亡患者死前血浆内毒素检测结果,推测内毒素血症“预后”线波动在(325±166)pg/mL,血中浓度超过此值时,有可能导致死亡。有鉴于此,我们认为循环内毒素水平对于革兰阴性菌感染及其他脓毒并发症的监测具有一定的预警意义。连续动态观察其变化,对脓毒症、MODS的早期诊断、病程监测、防治方案的制订及预后评价可能有指导作用。
表6-7 血浆内毒素水平与MODS发生例数
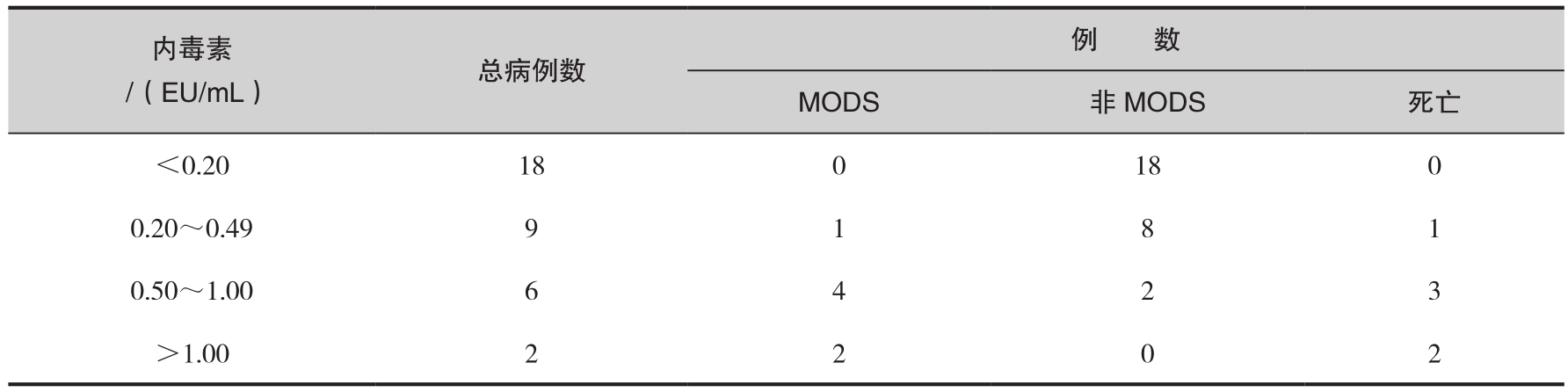
虽然采用抗菌药物治疗及其他强力支持性措施,但由于一般不能有效中和、灭活内毒素毒性或抑制其诱生的多种活性介质,因此难以从根本上解决内毒素血症的防治问题,严重烧伤脓毒性休克病死率至今仍高达70%以上。可以说,目前还缺乏有效的方法用于临床内毒素血症的治疗,成为提高大面积烧伤患者治愈率的一大障碍。现行的防治方案中,主要原则是及早全身应用抗革兰阴性菌的抗菌药物及正确处理烧伤创面。但有人报道,单纯使用敏感抗菌药物往往可迅速杀死细菌,导致短时间内大量的内毒素释放入血,在特定的情况下可能加重患者的临床毒血症反应。可见,对于内毒素血症的治疗仍存在许多矛盾,下面仅就有关的措施及其研究进展作一简要介绍,供读者参考。
早在20世纪60年代,Alexander等用多价铜绿假单胞菌疫苗(含抗脂多糖抗原及内毒素蛋白抗原)预防治疗烧伤患者,取得了一定的疗效,可提供部分保护作用。由于主动免疫抗体产生较慢,加之烧伤后多伴有免疫反应抑制,因此有人主张对于大面积烧伤患者采用高效价铜绿假单胞菌免疫球蛋白或免疫血浆进行被动免疫更为合适。这项措施可作为一项有效的辅助治疗,但存在来源困难、疗效仍不理想等问题,难以推广。为了克服上述缺点,近三十年来国内外学者致力于具有交叉保护作用的内毒素核心单克隆抗体的研制,早期大规模临床试用E5(来源于小鼠的IgM型抗J5类脂A单克隆抗体)抗体已显示出一定的疗效。在大量动物试验的基础上,人们对E5抗体进行了临床验证,表明该抗体是稳定、安全和易接受的。据报道,美国33个医疗中心使用E5单抗治疗468例可疑革兰阴性菌感染患者,结果证实治疗组(无论是否存在顽固性休克)与对照组相比,患者器官损害症状缓解或消失较为迅速,且非顽固性休克患者病死率显著下降,且过敏反应仅占1.6%,未出现明显的变态反应。在随后进行的另一组随机、对照、双盲临床试验中,E5单抗对革兰阴性菌感染患者却无明显疗效;但对于其中139例伴器官衰竭而无难治性休克的病例,E5单抗可以改善生存率。从目前的资料来看,还难以肯定E5单抗的临床治疗作用。参加临床试验的内毒素单抗制剂除E5外,还有一种来自人的IgM型抗体——HA-1A。一组543例脓毒症患者的临床试验资料显示,给予100mg HA-1A治疗组与对照组总病死率分别为39%、43%,无统计学差异。鉴于其中多数病例未发生革兰阴性细菌感染,他们进一步分析了197例培养证实菌血症患者对HA-1A治疗的反应,结果发现,HA-1A治疗明显降低并发革兰阴性菌脓毒症和脓毒性休克患者28d病死率。同时,HA-1A能够改善患者器官功能,治疗组与对照组器官功能恢复率分别占62%和42%。可见HA-1A抗体的作用可能主要限于伴有内毒素血症及革兰阴性菌脓毒症的病例。尽管关于内毒素拮抗剂在防治脓毒症中的有效性和确切临床价值仍有待于进行深入、周密的临床研究,但从目前已获得的大量资料来看,内毒素单抗制剂并不能明显改善脓毒症和脓毒性休克患者预后,因此,不宜过高期望抗内毒素抗体治疗的临床应用前景。
业已证明,多黏菌素B能通过化学作用结合内毒素的类脂A部分而中和或灭活其毒性,并且对铜绿假单胞菌、大肠埃希菌等常见革兰阴性杆菌有强大的抗菌活性。但由于这类抗菌药物不良反应大,尤其对肾的毒性而限制其临床广泛使用。为此,日本学者研制多黏菌素B固定纤维(PMX-F)治疗能选择性解除实验动物内毒素休克的毒性,从而显著地提高了动物存活率。近年来的临床应用报道有限,但初步证明有效。鉴于多黏菌素B的毒性反应,部分学者主张低于治疗量使用,在不产生明显的毒性作用的情况下,能减轻内毒素血症及提高烧伤患者的免疫功能,对改善患者预后是有利的。
杀菌-通透性增加蛋白(bacterial permeability increasing, BPI)是一种普遍存在于人及哺乳动物中性粒细胞嗜苯胺蓝颗粒中的蛋白质,相对分子质量约为55 000。BPI由两个活性部分组成: N -氨基末端具有杀菌活性,羧基末端起固定作用。BPI对内毒素的LPS分子具有高度亲和力,可有效阻止LPS激发的一系列免疫病理反应,对致死性内毒素血症具有保护性拮抗作用。同时,BPI对许多革兰阴性菌外膜有特异性结合能力,发挥其细胞毒作用,促使细菌外膜通透屏障破坏,导致不可逆性细胞损害及死亡。BPI强有力的中和内毒素及杀灭革兰阴性菌的双重特性表明,在革兰阴性菌脓毒症及MODS的治疗中可能具有一定的前景。绝大多数动物实验显示,完整BPI及其活性片段对小鼠、大鼠、家兔、猪、狒狒等革兰阴性菌感染或内毒素攻击均可产生良好的防护效应。Ⅰ期临床观察发现,rBPI 23 、rBPI 21 可显著抑制人体内毒素血症诱发的一系列免疫病理反应,证明它们是一种安全、高效的抗菌、中和内毒素制剂,故有人称之为“超级抗菌药物”。
BPI作为体内LBP-LPS相互作用的天然拮抗剂,其保护机制与直接中和内毒素、杀灭革兰阴性菌及竞争抑制LBP作用等多重效应有关。业已证明,BPI和LBP均为体内调控内毒素活性作用的蛋白,它们在序列上具有同源性及交叉免疫活性。BPI可与LBP竞争结合LPS分子,作为LBP-LPS复合物的拮抗剂存在。一般认为,LBP/CD14具有增敏机体内毒素的细胞毒性作用,介导LPS诱发的多种细胞因子的合成及释放;而BPI则可显著抑制或阻断炎症介质的产生及中性粒细胞补体受体的上调。BPI与LPS结合的能力比LBP强,rBPI 23 对LPS的亲和力大约为重组LBP的75倍,因此能竞争抑制LBP与类脂A结合。但当LBP/BPI比值增高时,BPI不足以抑制LPS的激活细胞效应;提高BPI的浓度则可以拮抗LBP的效应。严重烧(创)伤打击可广泛激活机体多种组织LBP mRNA表达,因此LBP/BPI比值明显增高,导致机体促炎和抗炎细胞因子比例发生失调,此时机体自身产生的BPI不足以抑制LPS的病理效应,因此给予外源性BPI以调节其平衡失调可能对改变机体的过度炎症反应状态及防止MODS的发生具有重要意义。并且BPI作为机体中性粒细胞中自然存在的防御性抗感染蛋白,是天然的LPS抑制物,具有其他外源性生物拮抗剂无可比拟的优点。研究表明,BPI及其活性片段均无明显毒性和免疫原性,在人体及不同动物模型中都能发挥强有力的结合及中和内毒素作用。据报道,大鼠、小鼠均可耐受高达10mg/kg的BPI而未产生毒性反应,1周后动物多个脏器功能与形态学未发现任何异常改变。因此,BPI作为临床抗感染治疗的重要辅助措施之一,可能具有潜在的应用价值。
Fine等使用口服非肠道吸收抗菌药物杀灭肠道细菌,减少内毒素的产生、吸收,取得了一定疗效。然而有些学者认为常规应用广谱抗菌药物并不能有效减少肠道内毒素和降低血中内毒素浓度,相反由于杀灭肠道细菌而引起内毒素的释放、吸收,可能使血中浓度升高。另一方面,临床上长时间应用广谱抗菌药物能使肠道微生态失调,存在诱发继发性感染的威胁。因此,有的学者提出针对肠道革兰阴性菌采取SDD的防治方法,能有效拮抗潜在性革兰阴性致病菌在宿主体内的定植,可能有助于减少肠源性内毒素的过量吸收,从而阻断内毒素血症的发生与发展。我们对应用SDD对烫伤动物全身性细胞免疫功能及预后的影响进行了探讨,结果证实,SDD组动物在给药3d内完全抑制革兰阴性肠杆菌的生长,其各段肠腔内游离内毒素含量较对照组下降99.5%以上,门、体循环内毒素水平随之显著降低,从而有效防止肠道细菌移位和内毒素血症的发生。经SDD处理动物可部分逆转严重烫伤所致细胞免疫抑制及巨噬细胞的活化,主要表现为脾细胞增殖反应及诱生IL-2活性显著升高,血清生物蝶呤水平及血浆TNF-α水平比对照组显著降低。同时,SDD组肠黏膜功能的标志酶——二胺氧化酶活性明显上升,动物5d存活率由63.3%提高至90.0%。说明预防性给予SDD能有效防治肠源性内毒素血症的发生与发展,其作用机制与减轻内毒素移位诱发机体细胞免疫抑制密切相关。但该方法能否改善患者的预后、降低病死率仍争议较大,有待于临床实践的进一步检验。
早期应用糖皮质激素、色甘酸钠等,可拮抗LPS活性,稳定粒细胞、肥大细胞释放血管活性物质,降低机体对内毒素的反应性,对缓解毒血症症状有一定作用。此外,一些学者提出抗细胞因子抗体干预,其中抗TNF-α抗体、IL-1受体拮抗剂(IL-1ra)最引人注目。阻断TNF-α可采用抗TNF-α抗体、可溶性TNF-α受体、TNF-α受体Fc段嵌合蛋白等多种方法。在注射活菌攻击后给予抗TNF抗体或TNF受体Fc段嵌合蛋白,研究结果显示出不一致性。早期治疗一般能获得较好的保护作用,延迟治疗在许多模型上缺乏保护作用,一般认为在内毒素攻击后延迟治疗不及攻击早期或预防性给药有效。尽管在动物脓毒性休克和致死性模型上拮抗TNF-α已取得令人鼓舞的结果,但抗TNF-α抗体在降低28d病死率中仍未被证明有效。对抗IL-1活性的药物有IL-1ra和可溶性IL-1受体。IL-1ra已应用于患有脓毒性休克患者的试验性治疗,多项临床Ⅲ期试验显示对患者预后均无明显影响,这与其他临床试验如用细胞因子或内毒素抑制剂治疗脓毒性休克结果相似。
总之,现代免疫治疗的目的是阻止机体由免疫中间产物所致炎症反应转变成为脓毒症状态。尽管有多种预防或治疗措施应用于脓毒症和MODS的防治,如抗内毒素单克隆抗体、抗TNF-α抗体、可溶性TNF受体、IL-1受体的拮抗剂等,但是临床大规模试验均未显示出完全有效的、可重复的、统计上有显著性意义的临床价值。这一残酷的教训使我们认识到,预防威胁生命器官损害最有效的方法是尽可能早期阻断或消除多种致病因素对宿主异常炎症反应和免疫功能的激活。一般而言,理想的增强机体防御能力的方法应当是预防创伤后全身炎症反应发展至不可逆转性自身损害效应。这种治疗一定要在损伤早期给药才能起到预防作用,它能保护多种靶细胞(如淋巴细胞、巨噬细胞、中性粒细胞和内皮细胞等),其作用方式可以防止宿主细胞过度激活和细胞损害。从我们目前对免疫机制的理解中可以看出,调节创伤后各种细胞间平衡失调最有效的方法可能是几种药物的联合应用。很明显,只有在机体遭受打击后及时通过对炎症、免疫细胞的直接干预以纠正其内环境紊乱状态,才能够有效预防脓毒症和MODS的进一步发生与发展。有人提出一种联合的免疫调理方案值得借鉴,主要包括:①急性损伤早期(≤72h)下调巨噬细胞和中性粒细胞的活性,上调淋巴细胞应答能力;②应用大剂量多价免疫球蛋白和可溶性补体受体,中和循环内、外毒素以防止巨噬细胞的过度活化;③重建细胞免疫功能,注射胸腺类激素、γ-IFN、粒细胞集落刺激因子来增强细胞介导的特异免疫反应以克服创伤后的免疫功能障碍。虽然在临床实际中采用免疫调理能否获得理想的效果受多方面因素的影响,但免疫治疗方案的本质不是被动、简单地“祛邪”,而是调动机体自身以“扶正”,即主动调节和恢复宿主防御反应,维护内环境平衡与稳定,真正发挥较全面的调理作用。
内毒素血症的防治仍是临床上一个棘手的难题,今后的方向可能是联合使用抗菌药物、拮抗内毒素与免疫调理治疗,并辅以强有力的支持措施。