
下载掌阅APP,畅读海量书库
立即打开



Hodgkin及Katz用钠学说(sodium theory)来解决上述事实与理论的分歧,提出动作电位期间,膜对Na + 的通透性 P Na ,远远超过 P K 和 P Cl ,从而有一股荷正电的Na + 骤然大量内流,使与 E K 接近的静息膜电位一跃变到与 E Na 相接近:

代入(表1-6-1)有关Na + 的数据, E Na 值相当于+55mV左右。Hodgkin与Katz等用蔗糖、葡萄糖或氯化胆碱代替外液中NaCl的方法所获得的结果与钠学说相当一致:[Na] o 减小时,超射也变小,[Na] o 降至正常值50%时,动作电位峰值减少约21mV,与Nernst方程推算结果相近。动作电位上升速率的减小也与钠学说相符。外液中[Na] o 降低50%,最大上升速率及 I i也降低50%。若此时全部离子流均由Na + 运载,Goldman-Hodgkin-Katz通量方程(公式1-7-8)所预计的电流下降58%,与观察结果极其相近。Kynes随后测出每次冲动 24 Na的净入量约为3.5μmol/cm 2 ,只相当于增加细胞内Na + 浓度的1/80 000,但却超过了膜电容充电到 E Na 水平所要求的数量,也肯定了钠学说。Hodgkin和Huxley进一步用电压钳制法定量地说明了钠学说的动力学。
顾名思义,电压钳是一种用电学方法控制膜电位水平,同时直接记录离子运动的方法。通过改变浸浴神经元细胞膜的溶液中的离子成分,可从所记录的
I
i中分辨出个别的离子成分,从而区分
I
i的载流离子与膜对该离子的通透性。根据
I
m=
I
i+
I
c=
I
i+
C
m
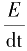 ,每次电压变动或跃迁时,会产生很小的瞬时
I
c,但电压一旦恒定之后
E
/dt=0,因而没有
I
c,
I
m可用
I
i表示,从而既可测定载流离子的种类,又可计算膜对有关离子的通透性。Hodgkin与Katz证明,当轴突去极化到0mV时,短暂的外向电容电流之后出现大的瞬时内向电流,最后再出现较大的持续外向电流。据推测,内向电流可能是早期
P
Na
升高期间Na
+
的内向运动产生的,后期的外向电流可能是K
+
的外向运动产生的。
,每次电压变动或跃迁时,会产生很小的瞬时
I
c,但电压一旦恒定之后
E
/dt=0,因而没有
I
c,
I
m可用
I
i表示,从而既可测定载流离子的种类,又可计算膜对有关离子的通透性。Hodgkin与Katz证明,当轴突去极化到0mV时,短暂的外向电容电流之后出现大的瞬时内向电流,最后再出现较大的持续外向电流。据推测,内向电流可能是早期
P
Na
升高期间Na
+
的内向运动产生的,后期的外向电流可能是K
+
的外向运动产生的。
Hodgkin和Huxley结合离子替换实验证明,以氯化胆碱代替海水中的NaCl时,电压钳制期间的早期内向电流消失,后期外向电流不受影响。他们结合反向电流测定更加证明,早期内向电流的载流离子是Na + :当电压钳制在 E Na 水平时(反向电位,reversal potential),不出现Na + 的净电流;如高于 E Na ,Na + 由内流变为外流,从膜内流向膜外(反向电流,reversal current)。证明载流离子最有说服力的方法是在电压钳制的基础上,将离子替换法与反向电流测定法相结合。在保持[Na] i 不变的条件下,改变[Na] o ,使其从[Na] 1 变到[Na] 2 时,Na + 平衡电位在理论上的变化值为:

与实测值符合,再次证明早期内向电流是由Na + 载流的。
利用乌贼巨轴突进行的早期实验中关于外向电流是由K + 载运的证据是间接的。外向晚电流的反向电位接近-77mV,与 E K 相近。虽然外向电流可由阳离子外流也可由阴内流形成,但以 42 K做示踪离子测定结果表明,K + 的外向总电流量与外向晚电流的积分值精确符合。电压钳制条件下证明:轴突内部灌流的K + 浓度降低时,外向电流变小,而且随着外液中K + 浓度的升高。外向晚电流的反向电位变得更正,与Nernst方程的定量一致。