
下载掌阅APP,畅读海量书库
立即打开



考虑到离子运动不受温度影响,不需能量供应,不受代谢影响;离子运动的速率和方向决定于膜两侧的电化学梯度;以及离子运动的速度极快和净通量很大等特点,多数学者倾向于用孔道或通道来解释离子运动及其对电压和时间的依从关系。近年来的突出发展表明,离子电导是在轴突膜上的一些散在而稀少的位点上发生的,这些位点的实质就是孔道。特别是在河豚毒(TTX)选择性阻断Na + 通透的启示下,对钠通道、钾通道以至钙通道的研究进展很快。通道的数目,单个通道的大小、构造、通透性和兴奋性等已成为各种新技术的研究目标。这些新技术主要有下述几种:①改变浸浴轴突的溶液成分并用电压钳技术测定动作电位发生过程中膜电流的变化,从而得到通道对各种离子的选择性并估计通道的口径;②测定伴随神经冲动出现的轴突光学或电学特性的变化,其中新近借以发现闸门电流(gating current)的新电压钳技术业已受到严格检验;③硫氢基活性因子效应,其结果可说明通道组成中蛋白质部分的存在;④利用放射性河豚毒和贝介毒(STX)在体内和溶液中结合钠通道成分,借以说明通道的数目和大小;⑤电镜技术的应用。
通道滤器(filter)指通道上的一种选择性滤器,它决定通道的选择性和通透性。通道对离子的选择性和通透性代表通道允许离子通过膜与区分不同离子的能力。Hodgkin学派曾用早期的钠通透性( P Na )和延迟的钾通透性( P K )说明动作电位发生过程中离子运动的变化。但是,当将轴突的轴浆几乎完全挤光并以不同成分的基质取代液灌注时,这两种通透性并不是绝对专一。在用乌贼巨轴突进行的电压钳实验中,Amstrong、Hille等测定各种基质取代液对早期Na + 流翻转电位的影响,并依恒电场方程,用有关公式计算钠通道对某一离子的通透性( P x )及其对Na + 通透性( P Na )的比值( P x / P Na )。例如 P K / P Na 的计算公式为:

式中 E r 为翻转电位的测定值, RT/F 为常数,膜外Na + 和膜内K + 的浓度([Na + ] o 、[K + ] i )已知,从而可以测出钠通道对K + 的通透程度。对于不能做轴突内灌注的无髓鞘纤维,在假定轴突内成分不变的情况下,可用其他一价阳离子代替Na + 做轴突外灌注,并依下式计算 P x / P Na :
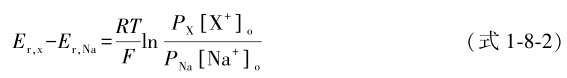
式中 E r,x 和 E r,Na 分别为X + 和Na + 灌注下测得的翻转电位,[X + ] o 和[Na + ] o 分别为待测离子和Na + 在轴突外灌注液中的浓度。在保持电压( E )恒定下,根据离子电流是离子通透系数与作用于离子的推力的乘积,也可用下式计算 P x /P Na :
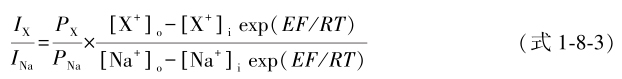
式中 I X 、 I Na 为电压恒定下X + 和Na + 灌注时测得的电流值,exp( EF/RT )为常量,其他有关离子的膜内外浓度为已知量。这些测定和计算也适用于钾通道通透性测定,但需以 P K 为标准测定 P X / P K 。测定表明:①钠、钾通道的离子通透性是相对的。它们除分别通透Na + 、K + 外,尚可通透其他无机离子和有机离子。钠通道还可通透5种无机离子和7种有机离子;钾通道还可通过3种离子。②钠通道的口径似比钾通道大,可以通透的最大的有机离子是氨基胍(0.37nm×0.59nm×0.76nm);可通透的一价无机阳离子顺序是Li + =Na + >K + >Rb + >Cs + >Tl + ; P K / P Na =0.83。钾通道只通透直径为0.26~0.3nm的阳离子,其顺序为Tl + >K + >Rb + >NH 4 + , P Na / P K 仅约0.01。③通透性的高低并不与离子大小绝对相关。直径最小的H + 不仅不通透,反而抑制其他可通透离子通透,有些二价离子通透极难(如Ca 2+ ,其 P Ca / P Na =0.01-0.1),与可通透的肼几乎等大的一些甲基化合物不能通透。
针对上述结果,Hille曾提出:钠通道是物理性孔道,其最窄部分的横截面为0.3nm×0.5nm,该窄区由8个氧原子环绕,其中包括一个带负电荷、pKa为5.2的离子化氧酸(ionized oxygen acid)。根据这个模型,比氨基胍大的有机离子所以不通透纯系几何学的原因;肼虽不小于0.3nm×0.5nm,但因其可与窄区氧原子在不同方位上形成许多氢键,因而可以通透,而形成氢键能力较弱的胍则很难通透。鉴于钠通道对有关离子的通透顺序与第XI序列高场强位点的结合顺序非常一致,故通道上负电荷可能具有较高的场强,因而能接受部分水合的离子。可通透的阳离子在通过窄区时与负性基结合,暂时失去其水合的一些水分子。离子直径为0.19nm的Na + 对负性基的离子引力比直径为0.266nm的K + 大,故Na + 通透易于K + 。由于负性基的负电荷在pH低于5.2时可能丧失,从而降低其对阳离子的引力,因而H + 不但不通透,甚至抑制其他离子通透。因此,阳离子通透钠通道的难易程度不仅与离子大小有关,而且与离子形成氢键的能力以及通道微环境的pH等因素有关。
离子通道除具有与通道滤器有关的选择性和通透性外,还具有兴奋性。这一特性主要指通道对膜电位变化发生反应的能力。这种能力与通道的电场感受器有关,后者又与启闭通道的闸门机构耦联。通道闸门通常被假设为只有开和关两种状态——兴奋时开,静息时关;因而离子通透性的变化显然取决于通道开放的数目。一度认为Ca 2+ 全或无地控制通道启闭,但实验表明,Ca 2+ 以及所有轴突内固有的可溶性小分子和离子,都与通道启闭控制无关。这一结果还提示,通道启闭既不需要Ca 2+ 等可溶性因子的辅助,也不需要代谢能供应,而是某种完全受制于膜内的内在过程。这种内在过程据认为就是当初Hodgkin学派所说的 P Na 或 P K 的激活和失活过程。许多实验证明这两个过程的存在及其各自的独立性。一种美洲东南部栖生的蝎的毒素能选择性地阻断激活过程,而另一种北非沙漠中生长的蝎的毒素则抑制失活过程,使通道持续开放达数百毫秒之久。海葵轴突外灌注、氟化物或以碘化物轴突内灌注均可引起类似的失活抑制效应。轴突内灌注蛋白分解酶,几乎使失活过程完全消失,钠通道持续地处于开放或激活状态。作用点为肽链中精氨酸的碱性蛋白酶以及对精氨酸有专一破坏性的乙二醛,均能阻断失活而不影响激活。这提示精氨酸在控制失活过程上可能具有重要作用。由于北非蝎毒能与钠通道发生1∶1的结合反应以及许多阳离子对这种结合具有竞争性,通道中也许存在能与北非蝎毒结合的、带负电的位点。这种位点一旦被有关毒素结合,通道即失去由开到关的能力。
尽管Tasaki曾对
g
Na
=
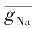 ·
m
3
h
,
g
K
=
·
m
3
h
,
g
K
=
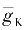 ·
n
4
g
Na+
=
·
n
4
g
Na+
=
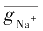 ·
m
3
h
的含义持疑,但人们还是从Hodgkin学派的论述中找到了这样的说明:
m
3
和
n
4
分别相当于3个和4个带电粒子运动到膜内某一正确位置,从而引起有关通道开放的概率;另一带电粒子运动到膜内某一正确位置,引起通道关闭的概率为(1-
h
),于是
h
代表该粒子未运动到正确位置的概率。这种去极时膜内有关粒子的运动规律,不仅证明是正确的,而且还被形象地比喻成3个“激活闸门”或“
m
闸门”(
m
3
)和“失活闸门”或“
h
闸门”(
h
)。
·
m
3
h
的含义持疑,但人们还是从Hodgkin学派的论述中找到了这样的说明:
m
3
和
n
4
分别相当于3个和4个带电粒子运动到膜内某一正确位置,从而引起有关通道开放的概率;另一带电粒子运动到膜内某一正确位置,引起通道关闭的概率为(1-
h
),于是
h
代表该粒子未运动到正确位置的概率。这种去极时膜内有关粒子的运动规律,不仅证明是正确的,而且还被形象地比喻成3个“激活闸门”或“
m
闸门”(
m
3
)和“失活闸门”或“
h
闸门”(
h
)。
在理论上,不论是带电粒子或闸门,它们的运动或开关均需有相应的能量对之做功。根据热力学分析,只有靠膜内电场通过膜来对闸门做功或使粒子运动才得以实现。如此,在离子电流出现之前,即应出现一种所谓闸门电流。近年在海兔神经元上,除发现与钠通道有关的闸门电流外,还记录到一种与钙通道活动有关的闸门电流。
有证据表明离子通道与镶嵌在轴突膜脂质双分子层中的蛋白质有关。许多重金属(如汞、银、铜、铀、镉等)离子以及选择性较强的硫氢基活性因子(NEM、PDMB等)均能降低轴突膜电兴奋性,其作用强度与其同硫化物生成不溶性盐的能力成比例,其效应又可被半胱氨酸或谷胱甘肽所阻止或翻转。这提示钠通道含有硫氢基并至少一部分是由蛋白质组成的。标本经清洁剂处理后Na + 电流能被蛋白酶、磷脂酶或胰蛋白酶降低的事实提示,通道蛋白质可能由磷脂所包裹。轴突内灌注蛋白酶和轴突外灌注北非蝎毒均能阻断失活过程的观察提示,通道蛋白质可能贯穿整个轴突膜。清洁剂涂层的TTX结合分子的沉降速率快和直径长的事实还指示,通道蛋白质是一个又大又长的分子,其分子量约为240 000Da。
除了不同的离子选择性和不同的电压与时间依从性之外,药理学的证据也提示钠、钾通道是彼此独立存在的。分子量不太大的有机阳离子——TTX和STX轴突外灌注能可逆地、选择性阻断Na + 电流,而不影响K + 电流;轴突内灌注四乙基胺(TEA)或其他季胺类离子只可逆地选择性地阻断K + 电流,而不影响Na + 电流。TTX与轴突标本呈1∶1结合反应的事实进一步提示,钠通道本身也是一个个单独存在的,因此,TTX滴定可测定单位膜表面积上钠通道的最高值。 3 H-TTX滴定的结果表明,海鳞鱼嗅神经的钠通道密度为2.5个/μm 2 ,大红虾腿神经为16个/μm 2 ,兔迷走神经为27个/μm 2 ,乌贼巨轴突为200~500个/μm 2 。通道密度有如此大的差别,据认为与轴突的直径大小有关,如乌贼巨轴突的直径为500~900μm,而海鳞鱼轴突仅约0.2μm。在轴突膜组分甚至该组分的可溶性蛋白质部分也测到类似的数据。这些数据虽很悬殊,但钠通道在轴突膜上的分布极其稀疏的情况显然可见。16个/μm 2 的通道密度意味着各通道之间的平均距离为250nm。
人们对钾通道的研究尚少。据设想,钾通道的最窄直径约为0.3nm,也由氧原子参与构成并具少量负电荷。由于通道口较窄,钾通道要求较完全脱水的离子。钾通道内侧口较大,足够四乙基胺进入并阻滞该通道活动。钾通道在乌贼巨轴突上的数目可能比钠通道多5~100倍,但钾通量仅为每微秒1个离子。钾通道的
n
4
闸门靠近内口,而其选择性滤器则靠近外口。钙通道分布虽较广,但人们对之所知甚少。钙通道可通透Ca
2+
、Ba
2+
和Sr
2+
,但不通透Mg
2+
。Mn
2+
、Co
2+
和其他重金属可阻断钙通道。钙通道的电导动力学参数曾提出过为
A
3
B
,与钠通道的
m
3
h
相似,但其激活与失活的时间常数是
g
Na
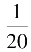 ~
~
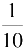 ,其开放的时程几乎与
g
K
相似,但不被TEA阻断。乌贼巨轴突神经冲动发生过程中,除Na
+
大量内流外,还有少量Ca
2+
内流。这种Ca
2+
内流虽不影响正常轴突的电反应,但轴突确有钙通道。钙通道多见于较慢的,常常是分级反应的细胞。
,其开放的时程几乎与
g
K
相似,但不被TEA阻断。乌贼巨轴突神经冲动发生过程中,除Na
+
大量内流外,还有少量Ca
2+
内流。这种Ca
2+
内流虽不影响正常轴突的电反应,但轴突确有钙通道。钙通道多见于较慢的,常常是分级反应的细胞。