
下载掌阅APP,畅读海量书库
立即打开



当感受器受到刺激或中枢发布命令时,在相应的传入或传出纤维上就可以检查出冲动。
神经冲动是沿着神经纤维传布的兴奋,是一种一旦产生就以恒定速度进行的波。
神经冲动并不是一股电流,它的本质是一个复杂的生物过程,是一种短暂的生物化学的变动。这种变动在神经纤维上产生和传导。
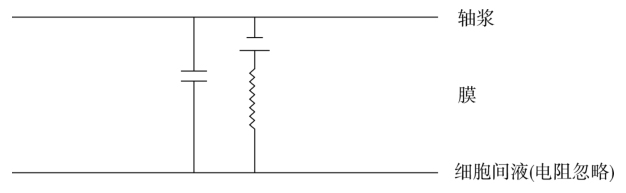
我们不妨将神经纤维看作一个柱状的管子,内部有导电的轴心(轴浆),外面有与纤维纵轴平行的导体(细胞间液),中间隔着具有高电阻和电容的膜,这种结构像一种电缆。
神经纤维中的轴浆是较稀的(0.3mol/L)的电离物质的溶液,具有较高的电阻率,再加上截面积很小,故轴浆对轴向电流的电阻很高,例如直径只有10μm的轴浆,1cm长的纵向电阻就有2.6亿Ω。轴浆内的电流只能在神经膜所包围的容积内流动。
与此相反,细胞间介质的容积则很大,电流流动的范围很广,因此细胞外液的电阻率虽然也高,有细胞内液的一半,但与轴浆的纵向电阻相比,细胞间液的总电阻却可以忽略不计。
膜的电学特性较为复杂,兼有电阻、电容和电池的特性。
离子只可以在有限程度上通过膜,因此膜有电阻性质。膜对于一种物质的透过性是该物质被膜两侧浓度差所推动而通过它的容易程度;而一种离子的膜电导(膜电阻的倒数)则决定于该离子被两侧电位差所推动时通过膜的容易程度。当推动力不变时,离子通过膜的速度取决于膜的结构。因此对一种离子而言,膜的透过性与膜的电导是膜的同一性质的两种不同的度量。
由于高电阻的膜将高度导电的内外液分开,或者说,由于膜的绝缘或分隔电荷的特性,膜本身又构成了一个电容器。虽然膜的电容性质并不改变通过膜的电流所引起的电压变化的方向,但是它对此电压变化的时间历程和空间分布确有显著的影响。亦即由于膜的电容与电阻特性,在外加电流作用下,膜电位随距离和时间的延长而呈指数式的衰减:电位改变的变慢是膜电容(储存电荷)所引起的后果;电位改变随距离而减小是膜电阻和轴浆电阻引起的后果。
膜的这种特性表明,神经冲动如要不衰减地传导,一定需要在冲动所经由的各处,以某种方式,加强所传递的信号。
综上所述,一小段轴浆和细胞间液的等效电路均是一个电阻器,一小段膜的等效电路则包括:①一个电容(代表膜的绝缘与储存电荷方面);②一个电阻(代表离子通过膜的有限能力);③一个电池,其电压为膜的稳定电压。这一小段轴突的整个等效电路可以表示为:
将相邻近的小段轴突的等效电路连接在一起,构成全轴突的等效电路。这种电学性质对轴突内外的离子分布和运动具有显著影响。
轴突膜内的轴浆与膜外的细胞间液之间虽有相似之处,但其区别却更为显著。两者之间既有显著的浓度差,也有一定的电位差。
细胞间液的Na + 比轴浆多10多倍,而细胞内K + 则超过膜外达30几倍。以猫的运动神经元为例,细胞内主要分布K + 和A - ,而细胞外则主要分布Na + 和Cl - ,其Na + 膜内、外浓度分别为15mmol/L和150mmol/L,K + 为150mmol/L和5.5mmol/L。
这种浓度和电位的巨大差别出现在轴突薄膜的两侧,因而有理由认为此膜对维持这种差别起着重要作用。
膜的这种作用表现在:①膜的类脂质与蛋白质交替排列的结构可能形成离子扩散障碍,例如同通过同等厚的水相比,K + 通过膜的速度为通过水的速度的1/10 000 000,同时膜对不同离子还有着不同的透过性或电导,例如静息时K + 的透过性比Na + 大约50倍。②轴突利用其代谢提供的能量主动地逆着浓度差将Na + 驱出膜外,将K + 驱入膜内,此即所谓的“Na + -K + 泵”。
综上所述,在静息条件下,离子在膜内外的浓度差、电位差、膜结构以及Na + -K + 泵等四个因素左右着离子的分布与运动。
Na + 虽有强大的由膜外向膜内扩散的趋势,但因膜对Na + 的透过性很小,少量漏进的Na + 又可被Na + -K + 泵驱出膜外,结果使Na + 的内流与外流速率相等,因而膜内Na + 始终保持低值。
相反,静息状态下,膜对K + 有较大的透过性,因而K + 顺着浓度差向膜外扩散。一部分K + 外流后,增加膜外的正电荷同时,膜内不能通过的有机负离子(A - ),则在膜的内表面牵制已经外流的K + ,阻止其远离。与此同时,随着K + 的外流,膜外正电荷渐渐增多而不利于K + 的继续外流,再加上Na + -K + 泵的主动活动,也使K + 向膜内运动。
结果,K + 的外流与内流的两种趋势几乎相等,但又不是正好相互抵消,而是略有扩散出细胞的趋势,因而由于K + 的运动,实际上给膜电容器充电,使膜的内外表面分别积聚了相反的电荷,形成外正内负的所谓极化状态,并具有稳定的电位差(膜电位)。
膜的兴奋性质,首先表现在快速的去极化(膜电位降低)使膜对Na + 的透过性增高或使Na + 载体激活,其次表现在Na + 的透过性增高或Na + 载体的激活是短暂的,常在几毫秒之内迅速降低或失活。
当刺激作用于轴突时,受刺激的部位极化减弱,膜电位降低(部分去极化)。一旦膜电位降到临界水平或阈限水平,即使Na + 载体激活,膜对Na + 的透过性急剧增高(约比静息时大500倍),于是Na + 顺着浓度差由膜外大量进入膜内,使膜内正电荷增多,发生完全去极化和反极化(膜内为正,膜外为负)的状态。
这种过程具有自生性或再生性。膜电位降到阈水平,使膜对Na + 的透过性增高,产生膜的更大的去极化,后者又进一步使Na + 载体激活,使Na + 的透过性更高,直至达到最大值。
然而,如上述,这种过程是极短暂的。紧接着的变化是Na + 载体失活,Na + 透过性逐渐降低,K + 透过性缓慢升高,再加上Na + -K + 泵的活动,结果使Na + 的内流减少,K + 的外流大量增加,直至K + 的外流量与先前Na + 的内流量在数量上大致相等,结果又恢复了外正内负的极化状态,即发生再极化或复极化。
再极化后,虽然极化和膜电位已恢复与静息时相同的状态,但膜内外的离子分布却与静息时不同。膜内有多余的Na + ,膜外有多余的K + ,于是再依靠Na + -K + 泵的积极活动,使K + 内流,Na + 外流,恢复静息时的离子分布。
如上所述,主要由于膜的兴奋性质的变化,Na + 载体的激活和Na + 的运动,产生了轴突的去极化,发生冲动或兴奋。Na + 一次一次地变动,去极化一次一次地出现,神经冲动即一次一次地产生。
神经纤维的电缆性质与兴奋性质间的相互作用,引起神经冲动的传播。
当膜的活动区去极化后,在轴突外表面的活动区与静息区之间出现电位差。这种电位差使电流从静息极化区,经过细胞间液,进入相邻的活动区。在这里,电荷通过膜流入轴浆,并经过轴浆,经由不活动区的膜流回,形成局部电流。
当这种局部电流这样流动的时候,非活动区的电荷减少,电压降低。若此局部电流流动的时间长到足以使不活动区的去极化达到阈限时,原来的静息区的Na + 载体即被激活,膜对Na + 的透过性急剧增高,结果此区也发生去极化和反极化,于是原来的不活动区就变成了活动区。
这种过程相继地在膜上的每一点依次重复,冲动就以恒定的速度和恒定的持续期,像波一样,沿着轴突的全长移动开来。人们将这种性质叫做传导或自行传播。神经纤维上的这种传播的变动就是神经冲动。活动区前面的局部电流就是使神经冲动相继出现,使不活动区兴奋的手段,因而局部电流事实上是去极化区传播或冲动传导的开路先锋。
神经冲动在传播过程中,在任何时刻,冲动或去极化区只占有神经纤维的一定空间(长度,即波长)。波长的数值,也即在同一时间内在神经纤维膜上呈负电部位的长度,可由动作电位的持续期与神经冲动的传导速度求知。不同纤维的波长不同,由数毫米到数厘米不等,最大的神经纤维冲动,波长可达6cm。
一次神经冲动在传导过程中,其大小和速度基本不变,是因为神经冲动的能量来自神经纤维各处的代谢供应,而不是来自刺激。
一条神经纤维上冲动的频率可有很大变化,由每秒几次到上千次。神经冲动频率的高低是神经元兴奋程度的表示。感受器引起的传入纤维的冲动的频率是感受器向中枢通报刺激强度的一种方式。
正常情况下,神经纤维上每秒内所传导的冲动可达100次。每秒经由Na + -K + 泵排出的Na + 量大约与每次冲动时Na + 的净内向通量相等。因此,当每秒传导100次冲动时,Na + 的内向通量比在静息时大100倍。为了保持兴奋性,细胞内部的Na + 必须保持很低的数值,因而持续活动时Na + -K + 泵的速度必须增加100倍。
上述过程足以解释无髓鞘神经纤维的传导,而有髓鞘的神经纤维的传导则牵涉到相同原理的略微不同的应用。
有髓鞘纤维的髓鞘中的髓磷脂是一种有效的绝缘物,它沿纤维全长每隔约2mm处以郎飞结断开。结间距离与纤维直径比较是极大的,约相当于100∶1。细胞膜被认为只是在这些结上才与细胞间液做有效的接触。有证据表明,局部电流是从一个郎飞结流到下一个郎飞结。因此神经冲动也就沿神经纤维从一个结到另一个结跳跃着,即跳跃传导。跳跃传导的一个结果是传导速度增快,因为使传播完成只需要把有限的几小块膜去极化就足够了。
此外,据认为,有髓鞘纤维的低膜电容与低轴浆电阻也是传导增快的重要因素。因为二者越低,则局部电流使邻近区域兴奋所需要的时间越短。
除Na + 、K + 等无机离子的作用外,Nachmansohn曾经主张,Ach的释放与破坏是神经冲动的必要步骤,并假定Ach的作用如下:①去极化作用使Ach从膜中固有被束缚的不活动形式中释放出来;②此Ach与一种“受体”蛋白质结合使膜对Na + 的透过性增高;③Ach与此受体蛋白质处于平衡状态,所以有些Ach是以自由形式存在的,很容易因高浓度Ach酶的存在而迅速水解(失活);④Ach的破坏,允许膜的再极化出现。
锋电位在刺激后立即出现,潜伏期很短(小于0.06ms),持续期也只有1个毫秒的几分之一,但在强度上却远远超过后电位。
锋电位按全或无原则活动。无论刺激多强,无论在神经纤维上传播多长一段距离,锋电位的大小不变。锋电位的大小只取决于纤维的种类和纤维当时所处的状态。由于锋电位的大小随纤维直径粗细而改变,因此锋电位与传导速度有关。直径粗,传导快的A类纤维比细纤维有较大的锋电位。
锋电位的持续期,在一定温度下是恒定的。与纤维的粗细无关。在A类纤维上,当单个冲动通过的时候,在任何瞬间大约有5cm长的神经纤维是处于活动状态的(即波长为5cm),在较粗的纤维上这一距离(波长)较短。
具有非常恒定特性的锋电位是动作电位的基本部分,亦即与神经冲动通过有关的这一部分。由于每引起一次冲动,总伴有一次锋电位;锋电位的传导速度与冲动的传导速度相等;以及当冲动被阻滞时,锋电位亦被阻滞,可见锋电位是神经冲动的必然表现,是传导性兴奋的标志。
后电位包括负后电位与正后电位,两者有某些共同的特点。①两者都是锋电位活动的后果,因而与锋电位有关;②与锋电位相比,两者均具有低振幅和长持续期;③与锋电位相比,两者均很不稳定,并与纤维的代谢状态和先前的经历有密切关系。增强神经代谢使后电位增加,但对锋电位无影响;缺氧、窒息和过敏时后电位逐渐减弱继而完全消失,但此时锋电位却依然存在。
由上可见,后电位与锋电位具有不同的生理意义。锋电位严格服从全或无原则,并与神经冲动相伴随;后电位则与神经纤维兴奋后的恢复过程密切相关,表示神经纤维对接受刺激已做好准备,或者如Gasser所说:“如果锋电位可以称为神经系统的信息传递者,则后电位可以认为是一种准备好能接受信息的指标。”
比较A、B、C类纤维动作电位的性质,虽然都具有基本上相同的组成成分,但在强度和持续期上却有着量的差别。
A类纤维:
A类纤维的动作电位振幅高、速度快、电刺激阈低,A类纤维中不同直径的动作电位表现出明显不同的高峰。另外锋电位的传导速度与振幅易受温度变化的影响。温度降低时振幅和速度均降低。
A类纤维的负后电位最高不超过锋电位的1/10,但它的持续期却长约15ms。刺激频率越高,持续期越长。A类纤维的正后电位的强度极低,大约只相当于锋电位的1/500,但是相对来说,它的持续期却是极长的,约为70ms或更长。
B类纤维:
B类纤维的动作电位与A类相比较,其特点在于:①锋电位的持续期长(1.2ms),相当于A类纤维的2倍,波形平滑,没有明显分离的高峰(即速度谱各处的纤维数相近);②没有明显的负后电位;③正后电位突出,几乎相当于锋电位的1/10。
C类纤维:
C类纤维的动作电位的特点为①锋电位持续期最长,约达2ms;②s.C具有显著的负后电位与正后电位,dr.C则没有负后电位,但显示出一种典型的、大的正后电位;③C类纤维的后电位不易受环境变化的影响;④电刺激阈值较高。
各类纤维动作电位各有其特点,其中亦有一些明显的规律性联系,如锋电位的振幅和传导速度与纤维直径成正比,而电刺激阈值则与纤维直径成反比。
外周神经干中含有不同数量的不同种类的纤维。由于它们的电刺激阈的不同,当给予神经干的电震,从阈限逐渐加大到最大强度时,其中的不同成分将以α、β、γ的顺序相继出现。
当给予神经干最大的电震,而使其中的所有纤维都兴奋,同时刺激电极与记录电极之间又有相当距离时,动作电位的形状通常是不规则的,表现出一系列在时间上先后不同的高峰。由于各类纤维传导速度的差异,高锋间的间隔,将随着传导距离的增加而增大。