
下载掌阅APP,畅读海量书库
立即打开



免疫原具有激活免疫系统产生免疫应答的能力,部分免疫原还可通过与免疫效应物质(抗体、效应性淋巴细胞)相互作用,进一步扩大或调节免疫应答强度。
免疫原性(immunogenicity)即指具备刺激机体产生固有免疫应答(炎症细胞因子效应)或适应性免疫应答(抗体或活化T、B细胞效应)的特性。免疫原的首要特性是具有免疫原性,是固有分子模式和抗原均具有的特性。
反应原性(reactionogenicity)指与免疫应答效应物质(抗体或活化的T、B细胞)相结合的能力,又称抗原性(antigenicity)。所有抗原具有反应原性;固有分子模式启动和激活固有免疫应答,但不能与炎症细胞因子等效应产物特异性结合,因此不具备反应原性(表3-2)。
表3-2 固有分子模式与抗原的比较

抗原同时具备免疫原性和反应原性,既能诱生特异性免疫应答,又能与免疫应答产物(抗体或活化T细胞)发生特异性结合。一些小分子化合物或药物,只能与免疫应答产物结合,却不具备刺激免疫应答产生的活性,被称为半抗原(hapten)。
半抗原只有与大分子蛋白质或多聚赖氨酸等载体交联和结合后,才成为完整的抗原。如青霉素降解产物青霉烯酸,本身没有免疫原性,但进入人体与组织蛋白结合成为完全抗原,诱导病理性IgE的产生;青霉烯酸再次与特异性IgE致敏的肥大细胞或嗜碱性粒细胞结合,导致Ⅰ型超敏反应的发生。
天然生物蛋白质大分子是良好的抗原,因其包含多个可诱导机体产生特异性抗体或激活T细胞的化学基团,这些化学基团被称为抗原表位。严格说来,淋巴细胞和抗体在免疫识别中的抗原特异性,是针对表位而不是完整的抗原分子。因此,抗原表位是蛋白质抗原分子诱导特异性免疫应答的最小结构与功能单位。
表位(epitope)又被称为抗原决定簇(antigenic determinant),是与T、B细胞抗原受体(TCR、BCR)及抗体特异性结合的基本结构单位,可由5~15个氨基酸残基、多糖残基或核苷酸组成。一个蛋白分子中能与抗体结合的抗原表位数目称为抗原结合价(antigenic valence)。天然抗原一般为大分子,含有多个不同抗原表位,为多价抗原;而半抗原仅有1个抗原表位,为单价抗原。
用生物蛋白质大分子免疫机体后,该蛋白质的多个不同表位均可选择相应的T细胞或B细胞克隆进行活化、增殖,进而生成免疫应答产物。因此天然蛋白质抗原免疫机体后,可得到针对不同抗原表位的多克隆抗体。
根据抗原表位氨基酸排列空间结构特点,分为线性表位(linear epitope)和构象表位(conformational epitope)。线性表位由连续的氨基酸构成,也称为序列表位(sequential epitope);构象表位则指不连续排列的若干氨基酸,在空间上彼此接近形成特定构象(图3-3)。T细胞不能识别游离的抗原表位,必须是在APC内加工并被自身MHC分子结合的线性表位。B细胞表面抗原受体(BCR)或者浆细胞分泌抗体识别的表位,既可以是线性表位,也可是构象表位,一般以构象表位居多。B细胞表位的氨基酸长度变化较大,为5~15个氨基酸残基,且多位于抗原表面,不需要APC呈递而直接激活B细胞(图3-4)。两类表位特性的比较如表3-3所示。
一般天然蛋白质抗原可同时存在T细胞表位和B细胞表位,即可同时活化T细胞和B细胞。而半抗原分子只有与蛋白质载体偶联后,才能诱导机体产生针对半抗原的抗体。其机制是:B细胞特异性识别半抗原,初步活化;而蛋白质载体中的T细胞表位,被B细胞或其他APC细胞呈递后活化CD4 + T细胞,通过载体表位使T-B细胞发生相互作用,T细胞通过细胞膜表面分子(如CD40L)及细胞因子(如IL-4)分泌,促进B细胞的活化和分化,最终分泌抗体。
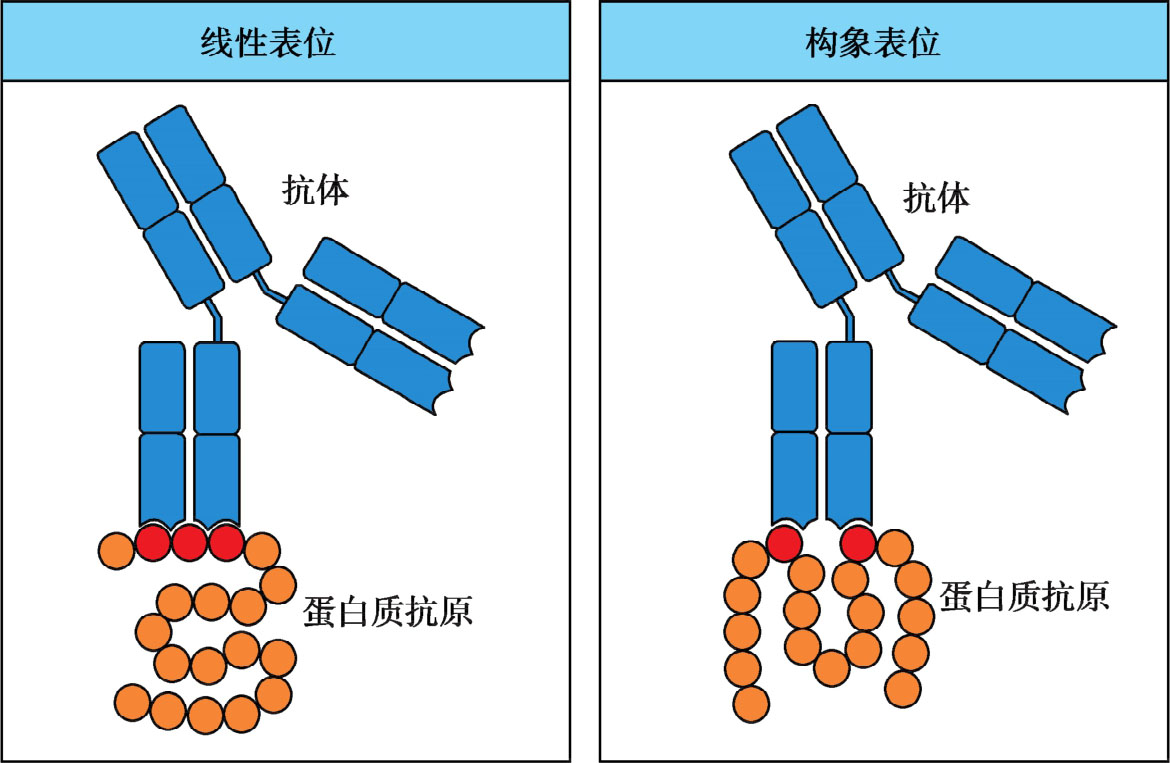
图3-3 线性表位与构象表位
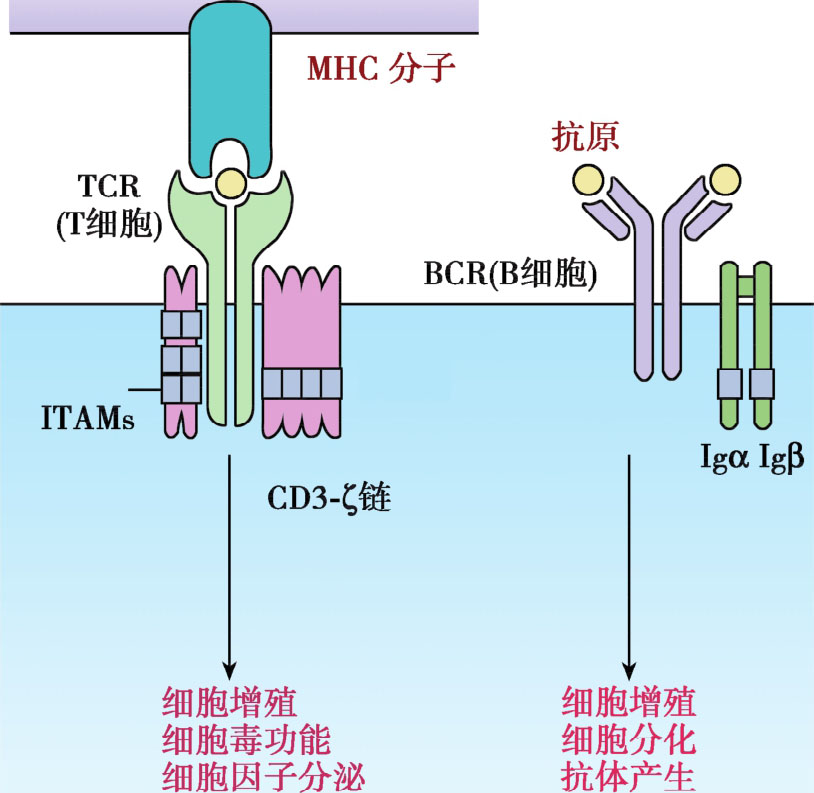
图3-4 TCR/BCR对抗原表位的识别
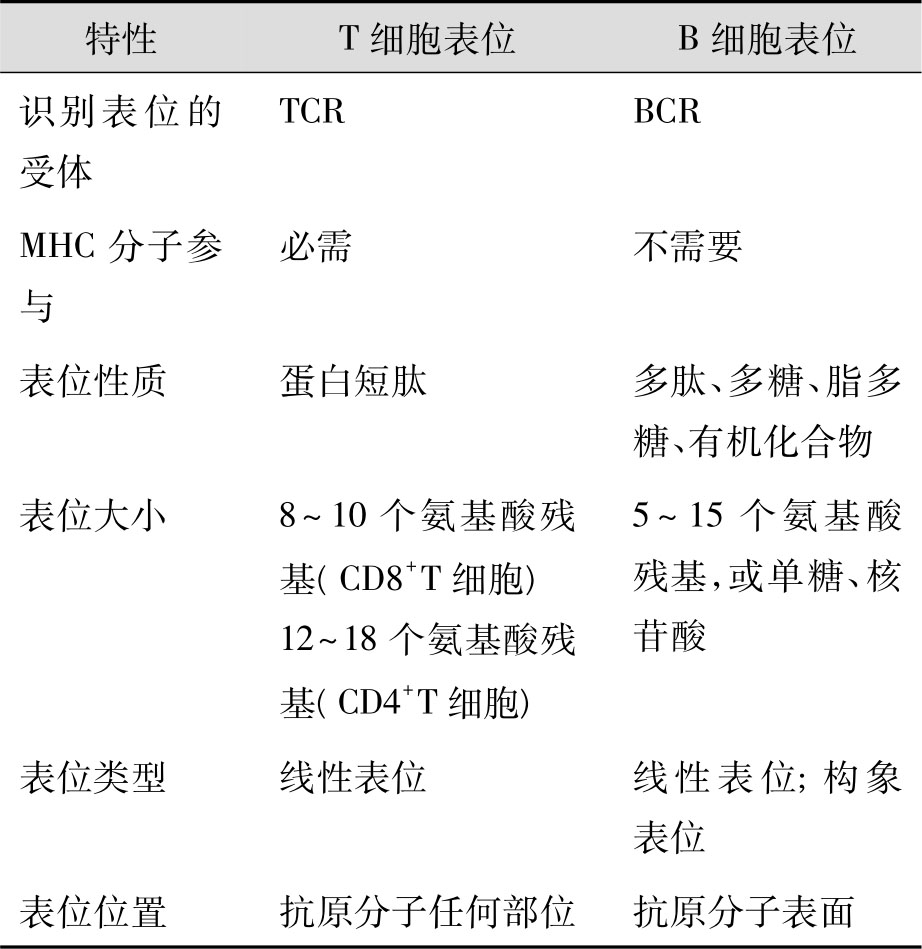
表3-3 T细胞表位与B细胞表位的特性
天然抗原分子含有多种抗原表位,而不同抗原分子可能含有相同或相似的抗原表位,称为共同抗原表位(common epitope)。因此,某些抗原诱导的特异性抗体或活化的淋巴细胞,不仅可与自身抗原表位特异性结合,还可与其他抗原中相同或相似的表位起反应,该反应称为交叉反应(cross-reaction)。