
下载掌阅APP,畅读海量书库
立即打开





现在我们要对前面讲过的内容做一点带注释性质的补充,但是这可能会使读者失去某些耐心,所以我想把它们变换成两个问题进行讨论。
第一个问题比较简单一点:早期人类是如何发现到达全球的那些迁徙路线的?前面提到过早亚洲人,也就是大约7万年前向东进发的那批现代智人,是一直沿着海岸行走的。从我们今天看来,这其实是一个很好的选择。因为原始人类的生存能力还极端低下,而在浅海附近,他们最容易获得动物蛋白的补充。但这并不意味着他们一开始就明白这一点。
在以采集、狩猎等攫取型经济来维持生计的时代,定居的大型人群将会很快因为四周生活资源的枯竭而陷入绝境。所以早期人类不得不分解成比较小的群体,在盲目地向外围世界不断离散和移动的过程中,去寻找各自的生存空间。在缺乏明确目标的漫游中,绝大多数群体由于遭遇这样那样的自然障碍,比如各种地理上的天险、瘟疫的袭击,以及其他的突发性灾难等,而归于灭亡。
只有一道接一道地通过了“瓶颈”压缩的少数幸运者群体,才得以继续生存。由这些幸运者群体无数次的偶然选择所连接起来的浪迹天涯的脚印,就是我们上面说到的“迁徙路线”。换句话说,这些所谓“路线”并不是出于早期人类的主动设计,而是他们以数不清的生命作为代价,去“试错”的结果。
第二个问题,有关人类起源和原始人群向全球迁徙的这么多信息,到底是从哪里来的?这些对生命科学研究者以外的其他人来说,至今还带有某种颠覆性的知识,主要来自分子生物学(也有人称之为“分子人类学”)在最近三四十年里面迅速发展的成果。在20世纪90年代以前的人类学教科书里,仅“人类”一个物种单独占据着动物遗传谱系里“人科”的位置,高居在“猩猩科”之上,但分子生物学完全改变了这一认识。原来属于“猩猩科”的黑猩猩、大猩猩等三个物种,今天已经完全被归入“人科”。
而与我们现在的讨论关系更密切的是,人们曾经普遍相信,现代人进入东亚的路径,是从西面先到达亚洲北部,然后再从北向南散布到东亚各地的。但是由分子生物学提供的信息,可以说是正好相反,现代人类是从南方进入东亚的。
分子生物学所关心的重大课题之一,就是在分子水平上考察和研究人类起源,以及不同人群间分化和融合的机理和历史。这里所谓分子水平,是针对男性人体细胞核内第23对染色体中的一条Y染色体上那一丝DNA而言。因为它就是由一个分子,或者更准确地说,是由一个长串复合分子构成的。如果把它拉长,可以有两米长,上面大约有6000万个碱基对,也就是基因片段。在父亲遗传给儿子的Y染色体的有些区段,基因片段的排列会因为Y染色体与来自母亲的X染色体的结合,而发生序列重组。但是Y染色体上有3000万个基因片段,是位于非重组区段的。
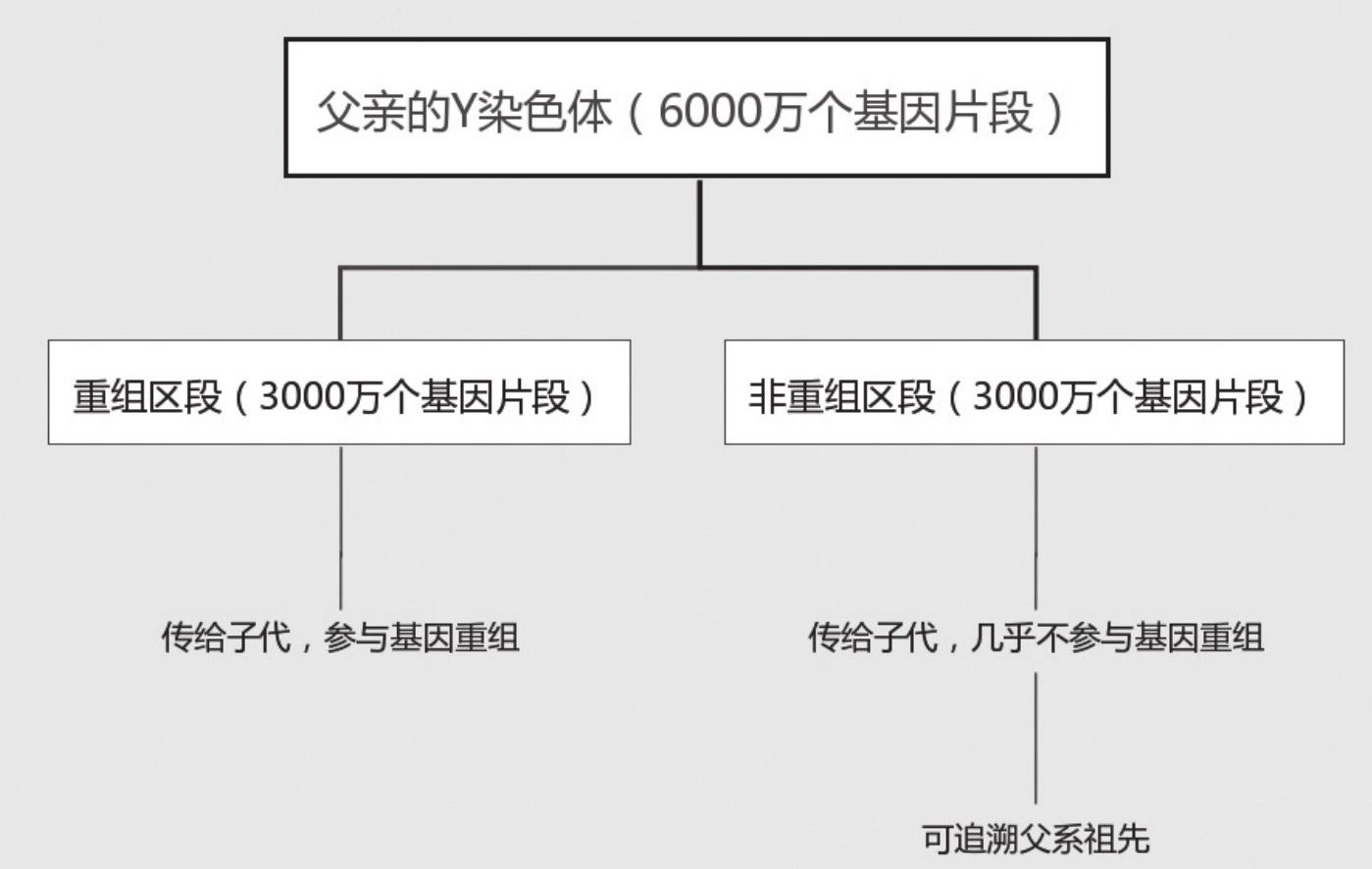
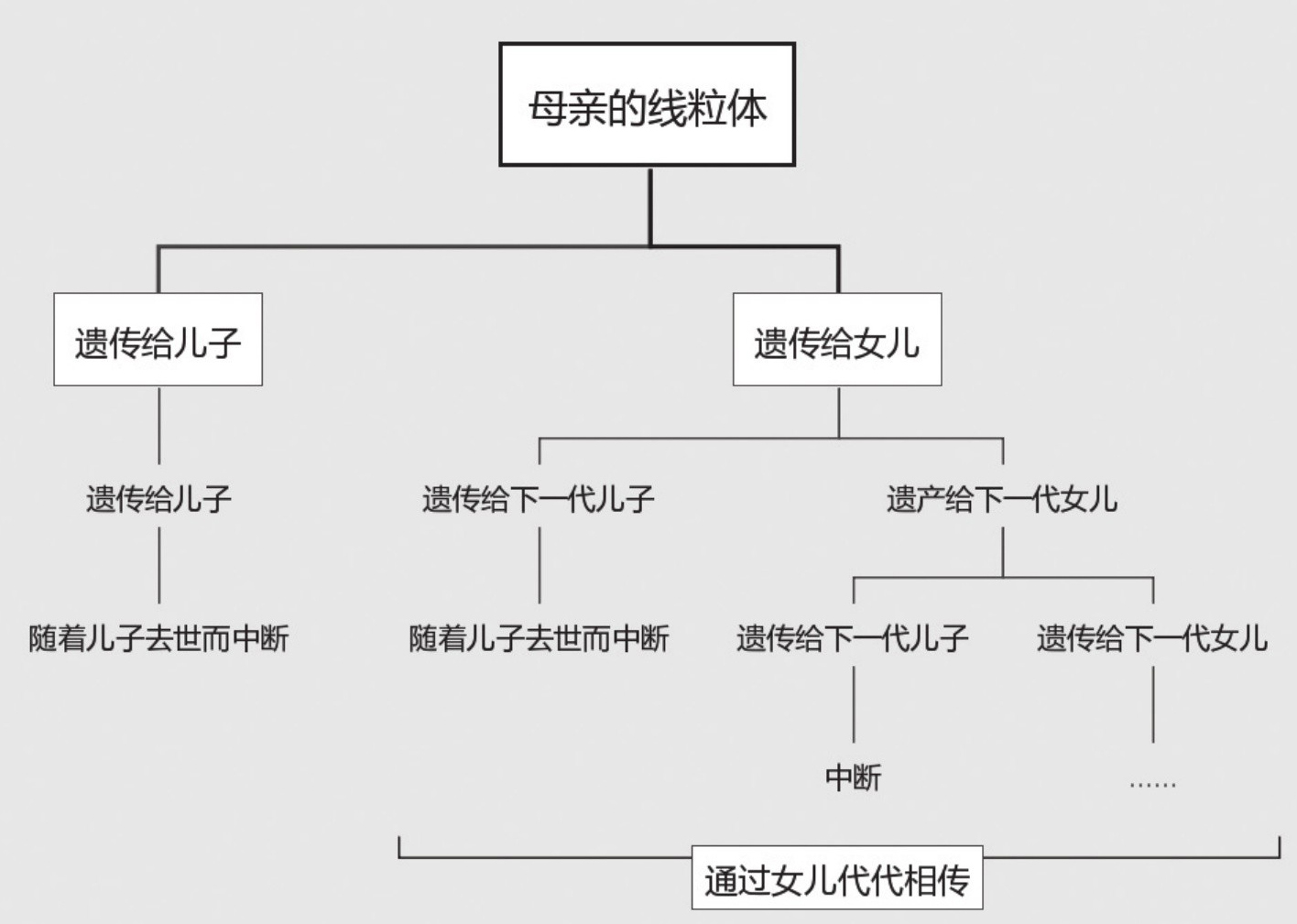
在从父亲到儿子的世代传递中,位于非重组区段的这3000万个基因片段,除每代平均有一个会发生随机突变外,都会以相同的编排序列,完整地由父代遗传给子代。而逐渐积累起来的基因突变也会一代接一代地遗传下去。因此如果样本齐全,根据这个区段上基因片段的排列,就不难把出于同一个父系的所有后裔成员编入一个血缘谱系之内。同样,人类细胞内的线粒体基因片段
 ,也可以由母亲完整地遗传给女儿,所以从线粒体DNA可以追溯一个女人的母系血缘关系。
,也可以由母亲完整地遗传给女儿,所以从线粒体DNA可以追溯一个女人的母系血缘关系。
有关人类DNA研究的突破,先是出现在对线粒体DNA的研究方面。20世纪70年代,全人类女性都出自一位非洲夏娃的发现,惊动了全世界。然而这项研究中的某些疏漏之处,引发了针对它的暴风骤雨一样的批评。从20世纪90年代起,随着检测技术的显著进步,不甘心的研究人员们重新振作起来,并且把考察对象更多地聚焦到Y染色体DNA上面。
现代人类是某一个非洲夏娃与某一个非洲亚当的集体后裔,对这一见解现在已经很少有行内人再会提出疑问了。一种似乎导向死胡同的研究路径,在经历一番柳暗花明的转折之后,重见天日。基因研究就是这方面的一个典范性的例证。
Y染色体DNA不仅可以用来追溯个体的父系血谱,也可以用来检测各个大型人群的独特遗传结构。不仅如此,对拥有各自特定遗传结构的不同人群之间分化和融合的相互关系,以及它的历史过程的探查,现在也有了可能。
早期人类从祖先群体中离散开来,四处游走,再加上他们因为一再遭遇各种自然与社会性的灾难,而发生群内人口严重耗损的所谓“瓶颈效应”
,都必然会导致祖先人群原先拥有的相当一部分根部基因变异在群体各个分支中丧失,尽管每个分离人群的下游基因变异可能在持续增加。因此,具有同祖亲缘联系的各人类群体的根部基因变异最丰富的所在地,便是那一系列同祖人群起源的地方。这就是论证现代人类起源于非洲的基本原理。研究线粒体DNA和常染色体DNA,对于揭示上述秘密,也可以发挥各自特殊的功用。
人身上的不同基因突变对后代产生的影响也非常不同。一方面,每个人群都会在一些特别时段生成若干对本群体父系遗传贡献非常之大的个体,从而把属于他们的基因突变传递给这个人群中占很大比例的后代成员。
另一方面,部分基因突变从人类生命史中被“抹去”,也不只发生在远古时代。没有生育或者没有生育男性后裔者的基因突变,也容易随世代的更替而最终丢失。这种不断发生的基因突变遗失,有效地抑制了被传递下来的基因变异多样化的规模。有些研究揭示出,生活在不同时期的对现代人有直接遗传贡献的人,也就是得以把属于他们自身的基因突变一直保留到现代人基因组中的个体,包括非洲夏娃和非洲亚当在内,总数不过8.6万而已。
我们现在有很多人对于没有后代会产生一种深刻的焦虑。但是如果你发现人类绝大多数的个体实际上都没有能留下自己的直系后裔,你还会受这一类的焦虑的干扰吗?
好了,现在可以结束这个可能已过于枯燥的科普话题,接着讲述现代人在进入中国境内后如何四处迁徙的基本情况了。
最早大约4万年前进入中国的早亚洲人,其实至少有两支。其中携带D单倍型的一支,应当是在四五万年前从中国云南与缅甸交界处的山谷走廊进入中国的。这支人群在青藏高原的羌族和藏语系民族中留下了很稀薄的遗传成分。东南亚以及印度洋东部安达曼群岛上的尼格罗陀人,则是尚未进入中国的D单倍型人群的后裔,虽然在其中有些人群里,这些遗传成分几乎已经完全丢失。他们是中国古代小说描写的身材矮小、肤色黝黑、短发卷曲,对主人忠心耿耿的“昆仑奴”的原型。
另一支携带C单倍群的早亚洲人,就像我们前面说过的,沿东海与黄海海岸线北上,在1.5万年前抵达黑龙江南北,成为今蒙古语和通古斯-满语各族共同祖先人群中最古老遗传成分的贡献者。蒙古语和通古斯-满语祖先人群的最早分化就发生在此之后。
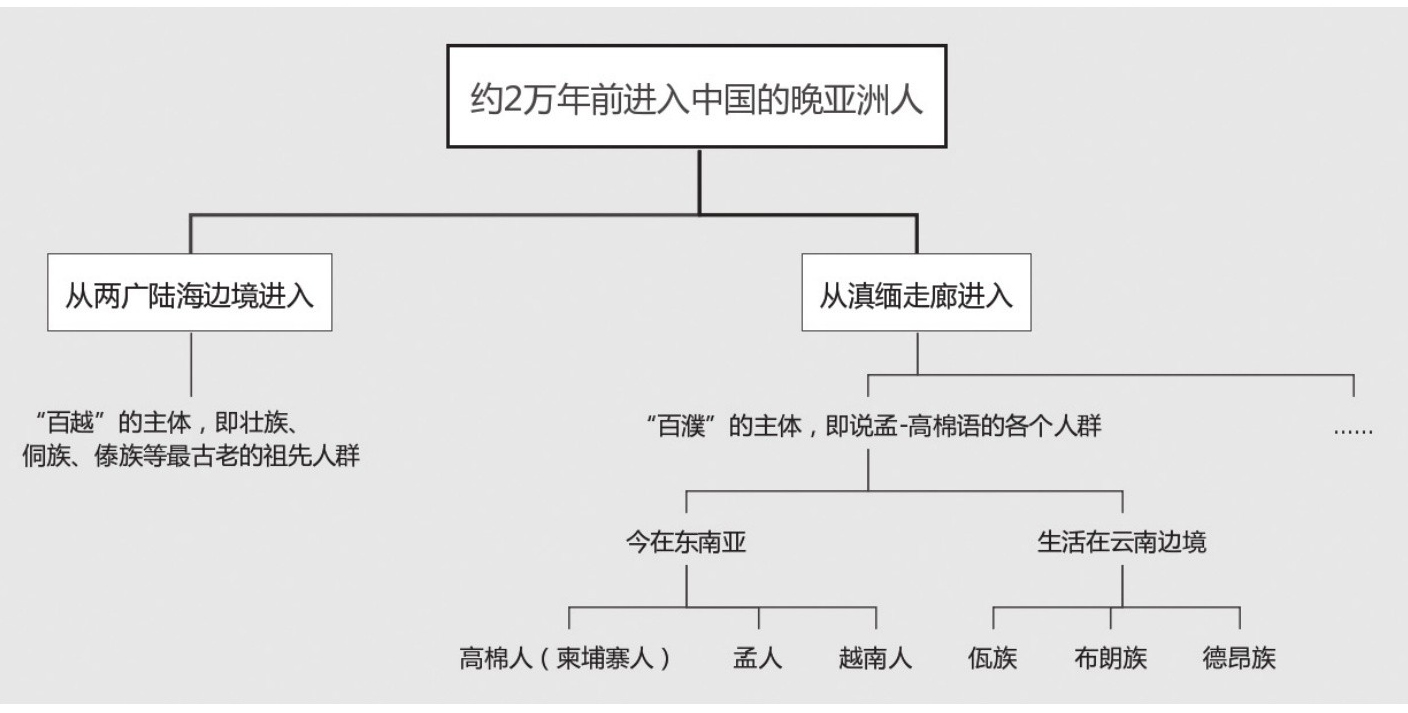
与早亚洲人一样,晚亚洲人也是分别从滇缅走廊和今天两广的陆海边界入境的,时间大约在2万年前。他们的染色体类型中最突出的是O型,还有少量的N型乃至更稀少的Q型和R型。今天靠近中国边界的东南亚邻国人口中有相当大的一部分,极可能源于已经进入中国,在人口膨胀后又重新溢出今中国版图的晚亚洲人。
从两广的陆海边境进来的那批晚亚洲人,是后来构成壮侗语系各民族,例如壮族、侗族、傣族、水族等族的最古老的祖先人群,他们恐怕是从上古直到秦汉记载中经常提到的“百越”的主要成分。直到东汉时,从今天的越南北部,北至杭州湾,仍然“百越杂处”,分布着骆越、夔越、夷越、瓯越、闽越、于越、南越、扬越、大越、山越等名目繁多的越人。他们未必都是同一种人,未必说同一种语言,但其中一定有相当大的部分属于壮侗语人群。
春秋后期,在江汉流域仍然有不少壮侗语人群的分布。两汉之际的刘向,在《说苑》这部书里采用两种方式记录了一首《越人歌》
,一种是它的汉语翻译,另一种方式则是借用汉字来记录《越人歌》的本来语音,类似于有些小朋友用“古特毛宁”四个字来记录英语“早上好”的发音。今天研究壮侗语的专家,可以毫无困难地借当代壮、侗语的知识,对汉代用汉字记音的文本从事试读。他们通过这个方式翻译出来的文本,所表达的意思与刘向记录的汉译本十分贴近。虽然刘向记录的译文可能因为深受汉代流行的楚辞风格影响,而已经变得“宛若楚辞”。
这个例证非常生动地表明,所谓“越人”中有很大一部分人口说的是属于壮侗语族的古代语言。他们沿海岸北上,走得非常之远。活动在上古中国北方的“东夷”中,可能还有他们的后裔,甚至主要就是由他们的后裔构成的。
如上所述,从两广陆海边境和从滇缅走廊进入中国的晚亚洲人,同样在南部中国后来的人口史和民族史中留下了不可磨灭的印记。但对华北来说,后一批人群,也就是从滇缅走廊北上的晚亚洲人的影响,相比而言就要大得多了。