
下载掌阅APP,畅读海量书库
立即打开



在本章第八节中我们谈到,在生命形成的初期,一些原核生物是靠氧化现成的还原性物质来获取能量的,例如氢气、硫化氢、氨都是还原性物质,它们的氧化会释放出生物所需要的能量,同时用氢原子来合成生物所需要的有机物。地球早期的大气中含有氢,火山爆发和海底热泉也会释放出硫化氢,不过随着时间推移,地球大气中的氢就越来越少了。氢虽然是宇宙中含量最丰富的元素,按原子数计算,宇宙中的氢占99%,在太阳系中,81.75%的分子也是氢,地球早期的大气中,也含有大量的氢,但是星际空间的物质密度是非常低的,每立方厘米大约只有一个微观粒子,相当于地球上的高真空。由于大气中的氢气很少有补充的来源,又由于它是最轻的气体,容易逃逸到外太空中去,再加上不断被细菌消耗,氢气的含量就越来越低。目前,氢气只占地球大气体积的千万分之五,少数能够产生氢气的细菌生产的氢数量也有限,所以氢气不可能作为地球上生物丰富可靠的能源。火山喷发和海底热泉释放的硫化氢数量也有限,所以硫化氢也不会是丰富和永久的能源。地球上的生物要大发展,就需要一种无处不在,而且永不枯竭的能源,这就是太阳光。地球轨道上的平均太阳辐射强度为1369瓦/平方米。从地球赤道周长约为四万公里来计算,地球获得的能量可以达到173000TW(太瓦)。地球在一小时内获得的太阳能,比人类一年内使用的能量还要多,而且可以持续数十亿年。
太阳辐射能的99%集中于波长为150 —7000 纳米的电磁波中,其中可见光区(390—700 纳米)占总辐射能的约50%,红外光区(波长大于700 纳米)占约43%,紫外光区(波长小于390 纳米)只占约7%(图2-28)。光辐射能量的大小与它的频率成正比,即E = hν,这里E是光子的能量,ν是它的频率,h为普朗克常数,为1.58 ×10 -34 卡·秒。所以光子的能量和它的频率成正比,而和波长(光速除以频率)成反比。波长越长,能量越低,因此紫外光的能量高于可见光,可见光的能量又高于红外线。
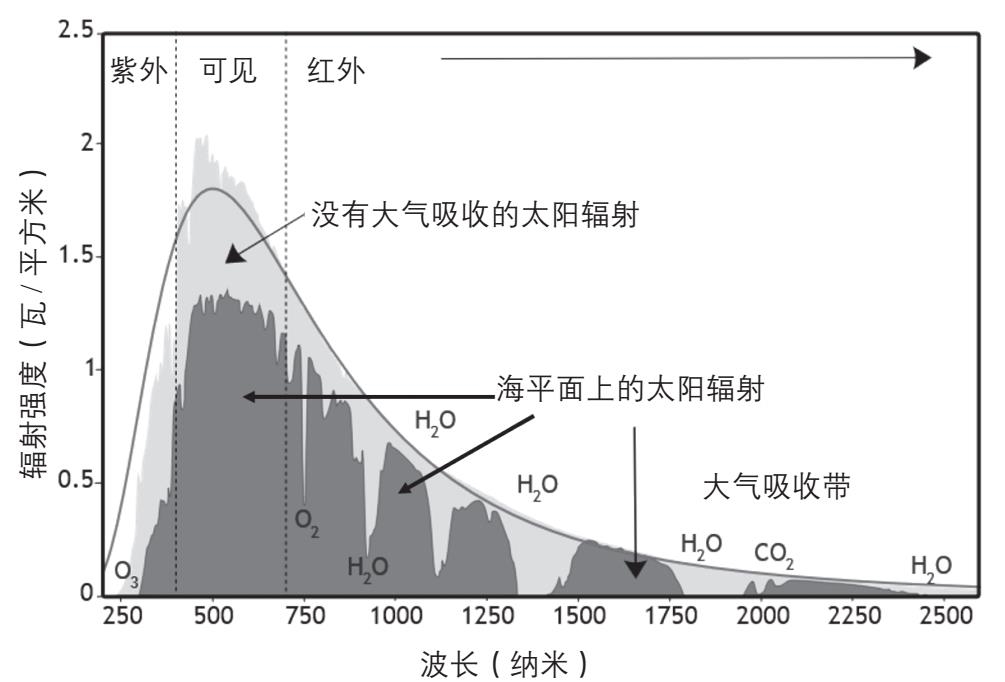
图2-28 地球上接收到的太阳辐射
从图2-28可以看见,臭氧(O 3 )对紫外线吸收很强,所以到达地面的紫外线大大减少;水和二氧化碳在红外辐射区间也有几个吸收带,但是它们对可见光的影响不大。
虽然地球上有丰富的阳光资源,但是要捕获太阳光中的能量,使它变为生物能够利用的形式,却并不容易。一是要有合适的分子来吸收光能,二是吸收的光子所含的能量必须合适。紫外线的能量太高,容易造成化学键的断裂,红外线的能量太低,只能增加分子的热运动,都不适合作为生物的能源,能够作为生物有效能源的,主要是可见光和波长接近可见光的红外线。
在吸收可用光能上,氨基酸、核苷酸、脂肪酸以及由它们组成的蛋白质、核酸和脂肪都派不上用场,因为这些分子含有的化学键大部分是单键(例如碳-碳单键和碳-氢单键),而单键吸收光能实现电子跃迁需要的能量比较多。例如甲烷的吸收峰在125 纳米,其他饱和烷烃的吸收峰在150 纳米左右,都在紫外区。
如果在碳原子之间形成双键,电子跃迁需要的能量就小一些,例如乙烯的吸收峰就在185 纳米。碳原子之间在形成双键时,如果双链之间再形成共轭双键(conjugated double bond,即被单键隔开的双键),电子跃迁需要的能量就更低,例如丁二烯(2个双键共轭)吸收峰就移到217 纳米。由于这样的共轭系统在生物的有效光吸收上扮演关键角色,我们在这里稍微多介绍一下其中的原理。
在第一章第一节中,我们已经介绍了碳原子之间以双键相连时,一个键是通过sp 2 杂化轨道形成的,另一个键是未改变的2p轨道彼此重叠形成的(见图1-1),我们把重叠的p轨道叫做π轨道,把π轨道中的电子叫做π电子。由于碳原子之间是通过双共价双键相连,它们之间的距离(即双键的长度)为13.4纳米,明显短于碳-碳单键的15.4纳米。如果有两个碳-碳双键,彼此被一条单键隔开,这两个双键的π轨道又可以彼此部分重叠,形成一个共同的大π轨道(图2-29),使得隔开两个双键的那条单键变短,从15.4纳米变为14.8纳米,成为部分双键,相当于2个双键和它们之间的一个弱一些的双键融合在一起,使电子活动的范围更大,跃迁需要的能量更低。这种π轨道可以彼此重叠的双键就叫做共轭双键。
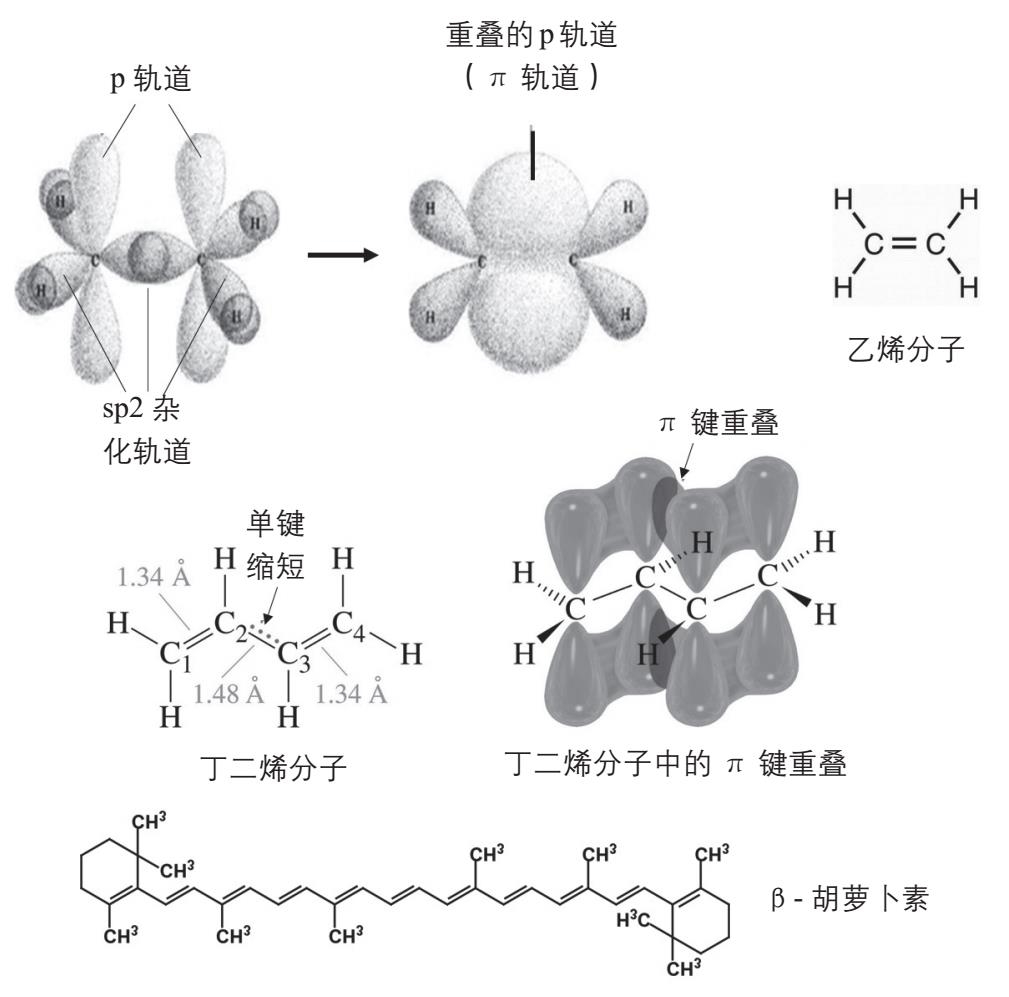
图2-29 碳碳双键和共轭双键。上图:两个碳原子以双键相连时,4个外层电子形成3个sp2杂化轨道和1个未变的p轨道。两个碳原子用1个sp2轨道和1个p轨道的重叠来形成双键。中图:在丁二烯分子中,两个双键的4个p轨道(即两个π轨道)可以重叠在一起,形成大的π轨道。下图:更多的双键可以形成更大的共轭系统,例如β-胡萝卜素
除了两个双键可以形成共轭双键,多个被单键隔开的双键也可以形成更大的共轭系统,即更大的π轨道,因为它们的π轨道也可以像丁二烯分子内的两个π轨道那样彼此重叠。参与共轭双键体系的双键数量越多,π轨道的范围越大,电子跃迁需要的能量越少。例如1,3,5-己三烯(3个双键共轭)的吸收峰移到285纳米,癸五烯(5个双键共轭)的吸收峰在335 纳米。虽然如此,这些化合物的吸收峰仍然在紫外区。要有效地捕获光能,需要更大的共轭系统,把吸收区域转移到可见光的范围内。一些分子由于含有巨大的共轭系统,可以吸收某些频率的可见光,我们能够看到没有被它们吸收的可见光的颜色,所以这些分子称为色素(pigment)。例如β-胡萝卜素(β-carotene)分子的共轭系统含有11个碳碳双键,而且都被单键隔开,它们就可以组成一个大的共轭系统(见图2-29下),吸收从紫色到绿色的可见光(400—550 纳米),我们看见它的颜色就是橙红色的。
地球上的生物主要利用两种色素分子来捕获光能,即由异戊二烯单位组成的线性大共轭分子(例如视黄醛和胡萝卜素)和由卟啉环组成的环状大共轭系统(例如各种叶绿素)。这也没有什么可奇怪的,因为生物都是尽可能将已经有的分子加以改造,让它们具有新的功能,而不是一切从头做起。聚异戊二烯链类型的结构在原核生物中早已存在,例如细菌呼吸链中的醌就含有聚异戊烯的长“尾巴”(见图2-33)。细菌的呼吸链中许多电子转运蛋白都含有血红素辅基,叫做细胞色素(cytochrome),例如细胞色素b、细胞色素c等等。而叶绿素的分子结构和血红素非常相似,都是由四个吡咯环(由4个碳原子和1个氮原子组成的环)通过次甲基( CH—)彼此相连组成的大环状结构(见后文及图2-31)。叶绿素和血红素的合成路线也相似,都是以谷氨酸为原料,经过琥珀酰辅酶A和氨基酮戊二酸(aminolevulinic acid)合成“胆色素原”,再由“脱氨酶”把胆色素原连成一个卟啉环。这一过程说明叶绿素和血红素有共同的分子起源。只要把血红素的合成路线做一些修改,就可以合成叶绿素。叶绿素有多种形式,被不同的光合生物所使用,但是它们的基本构造是相同的,即基本构造是卟啉环,功能也都是捕获光能。
根据色素分子受光照射后是否射出电子,生物利用光能有两种方式:第一种是色素分子不射出电子,而是利用光能直接把氢离子从膜的一边泵到另一边去,创造一个跨膜的氢离子梯度。这个氢离子梯度就像水库里面蓄的高位水,可以在流回膜的另一边时,带动ATP合成酶生成ATP(见本章第九节,原核生物“蓄水发电”)。有些原核生物就采用了这种方式,例如有些古菌是“嗜盐菌”,可以在饱和盐溶液中生活。其中的盐杆菌(halobacteria)细胞膜呈紫色,因为膜上含有一种蛋白质,叫细菌视紫红质(bacteriorhodopsin)。它有7个跨膜区段,在细胞膜内围成筒状,在筒的中间含有一个色素分子叫视黄醛(retinal),分子中含有多个异戊二烯单位。视黄醛含有由5个双键组成的大共轭系统,其吸收峰在357 纳米的紫外区,所以还不足以在可见光区域有吸收。但是当它上面的醛基与细菌视紫红质蛋白上216位的赖氨酸残基上的氨基结合后,吸收峰就被移到可见光中500—650 纳米的区域(绿色和黄色),其吸收峰在568 纳米,所以看上去为紫红色。它在受光照射时可以改变形状,从基本上是一根“直棍”变为“弯棍”。这个形状变化就把氢离子从蛋白质位于细胞质附近第96位上的天冬酰胺残基转移到位于膜另一边第86位的天冬酰胺残基上,再释放到细胞膜的外面。所以细菌的视紫红质可以直接利用光能来产生跨膜氢离子梯度(图2-30)。这个质子梯度就是能够储存的一种方式,可以用来合成ATP。
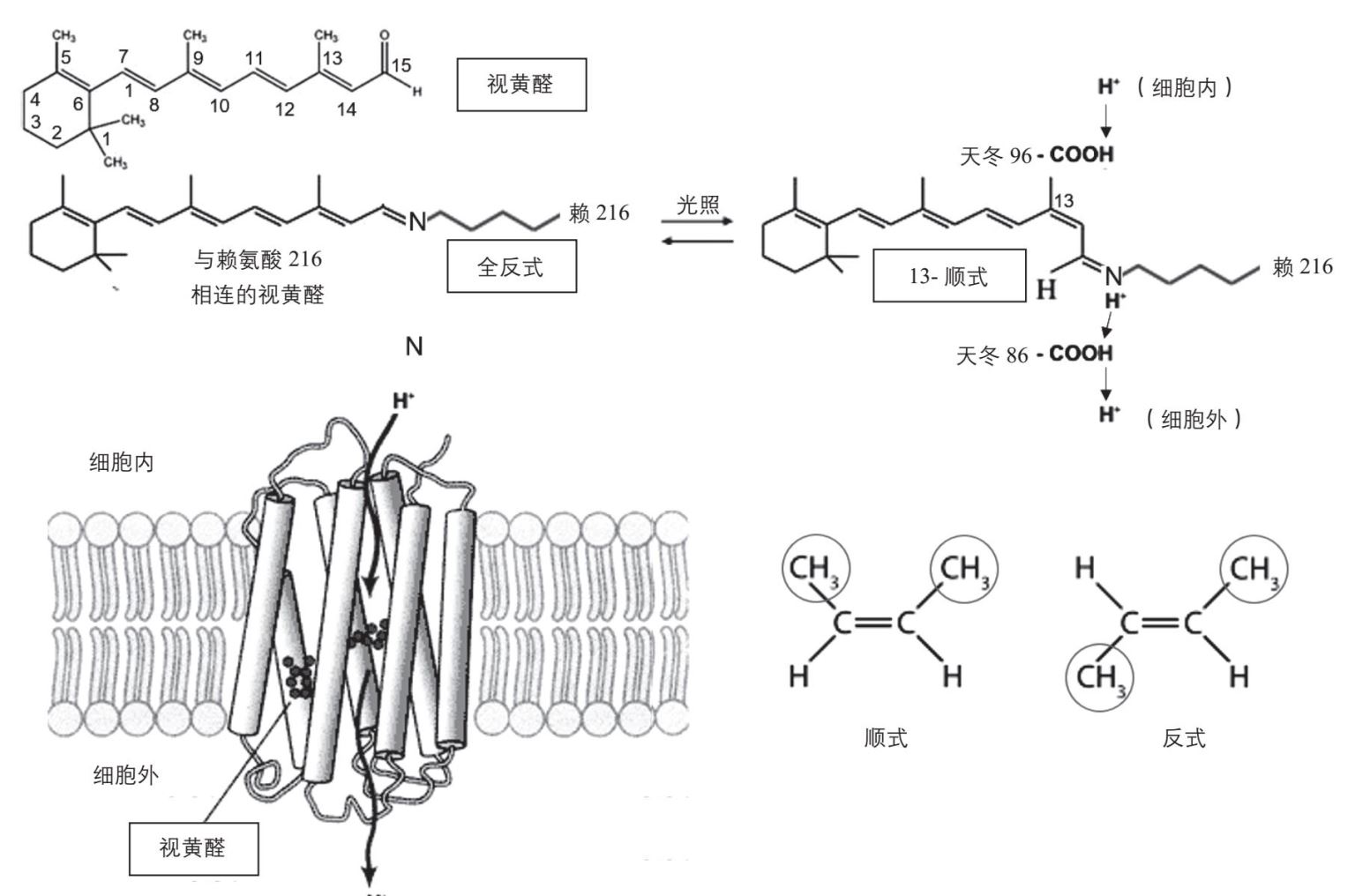
图2-30 细菌的视紫红质可以利用光能直接产生跨膜质子梯度
视黄醛的分子受光激发时形状从“直棍”变为“弯棍”,在化学上就叫从“全反式”变为“13-顺式”。要知道什么是反式和顺式,就需要回忆一下碳碳双键的性质(见图1-1)。两个碳原子以双键相连,好像两个塑料球被两根牙签穿在一起,是不能相对转动的。这两个碳原子余下的两根单键和双键也在一个平面上。如果这两个碳原子上各自又连有碳原子,就像在碳链中的情形,就有两种连接方法。一种是外接碳原子在双键的同侧,叫做顺式连接;另一种是外接碳原子在双键的对侧,叫做反式连接(见图2-30右下)。反式连接对碳链形状影响不大,碳链总的来说还是一条直链,但是用顺式连接时,碳链就必须在这里有一个“拐弯”。在没有光激发时,视黄醛分子内所有与双键的连接方式都是反式的,所以视黄醛分子像一根“直棍”。当受到光激发时,第13位碳原子处的双键就会从反式变为顺式,“直棍”也就变为“弯棍”。这个变化是可逆的,随后顺式视黄醛又会自己变回全反式,可以再次被光激发改变形状。视黄醛的这个性质非常有用,不仅被细菌用来直接建立跨膜质子梯度,还被动物用来获得光线中的信息。我们在第十二章第一节中,还将介绍视黄醛的视觉功能。
与蛋白质结合的视黄醛虽然可以把光能直接转化为跨膜质子梯度,继而合成ATP,给原核生物的有机合成提供能量,所以也可以属于广义光合作用的范畴,缺点是不能提供有机合成的氢原子,细菌还得从现成的有机物中获得氢原子,所以只能算作半个光合作用。如果还有一种机制可以同时将光能用于ATP合成和提供氢原子,生物就可以摆脱对现成有机物的依赖,自己从无机物合成有机物。这个任务是由叶绿素来完成的。
叶绿素的分子含有一个复杂的环状结构,叫卟啉环。卟啉环里面所有的双键都是彼此“共轭”的,即彼此都被一个单键隔开,因而形成一个巨大的共轭系统。不仅如此,卟啉环的中间还连有一个镁离子(见图2-31左下)。这个结构使得叶绿素在紫外和可见光区段都有吸收峰。不过叶绿素最早的功能也许还不是进行光合作用,而是结合在膜蛋白上,起保护DNA不受太阳光中紫外线的伤害作用。在地球大气中氧气出现以前,大气上层还没有臭氧层来阻挡紫外线。强烈的紫外辐射对于原核生物的DNA是很大的威胁,因为紫外辐射能够使DNA链断裂,碱基突变。既然所有的生物早就能够合成血红素,出现类似叶绿素那样的分子也不是难事。与膜蛋白结合的叶绿素不但能够吸收可见光,也能够吸收紫外线,相当于我们皮肤细胞里面的黑色素保护我们少受紫外线的伤害。
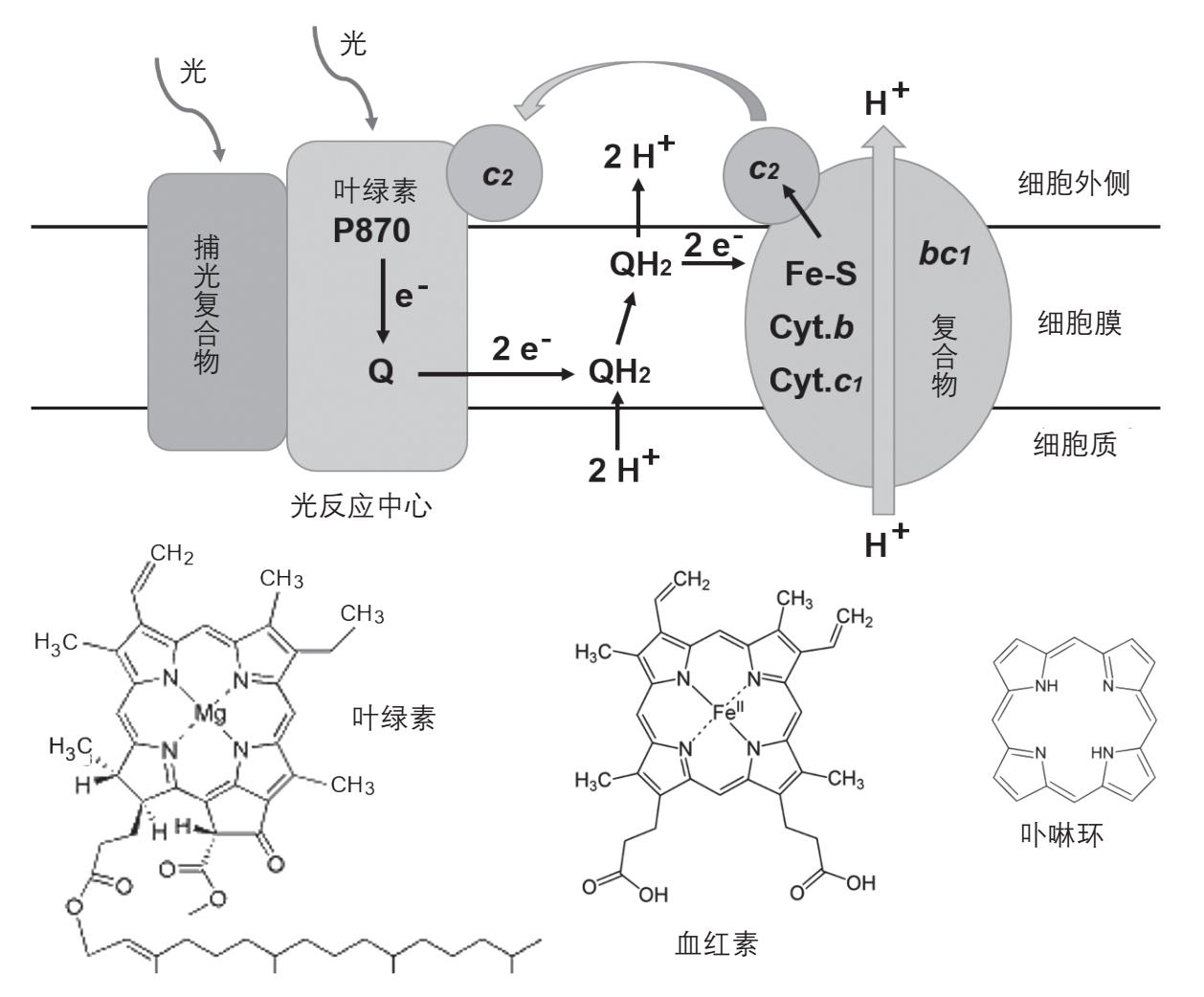
图2-31 上图为紫细菌的光反应回路。下图比较叶绿素和血红素的分子结构。它们都含有一个卟啉环,中心都含有一个金属离子,在叶绿素是镁离子,在血红素是铁离子
在叶绿素分子结构微调的过程中,一个变化发生了,就是叶绿素受到可见光激发时,可以射出一个电子。如果这个电子可以从细胞膜的一边转移到另一边,而丢失电子的叶绿素分子带一个正电,就可以发生膜中的电荷分离,类似于跨膜氢离子浓度,因为二者都有膜两边电荷不平衡的情形,也是储存能量的一种方式。如果有一个机制能够把这个电荷分离变成跨膜氢离子梯度,光能就可以用“水坝蓄水”的方式储存起来了。这个任务是由原核生物的光反应中心来完成的,是原核生物利用光能的第二种方式。下面我们以能够进行光合作用的原核生物紫细菌(purple bacteria)为例,看这个过程是如何完成的(图2-31)。
紫细菌的光反应中心含有一对叶绿素分子,它们结合在蛋白质上,位置靠近细胞膜的外侧。受到近红外光激发时(吸收峰在870 纳米),这一对叶绿素分子射出一个电子,经过几个中间步骤将电子送到细胞膜的内侧,同时在叶绿素分子上留下一个正电荷。转移到细胞膜内侧的电子被一种叫做醌(quinone,简写为Q)的分子所接收。不同生物所使用的醌类物质在结构上有一些差别,在紫细菌中为泛醌(ubiquinone),在蓝细菌和绿色植物中为质体醌(plastoquinone,其结构见图2-33下),但是它们的功能是相同的。当反应中心受激发两次时,醌分子就接受两个电子,再和内侧细胞质中的两个氢离子结合,形成与醌相连的两个氢原子,让氧化型的醌Q变成还原型的QH 2 。这样就在细胞膜的内侧消耗两个氢离子。QH 2 是高度脂溶性的分子,而且不带电,从蛋白复合物上脱离后,可以在细胞膜内的脂质环境中游动。当它到达细胞膜的外侧时,会遇到另一个蛋白复合物叫做 bc 1 复合物, bc 1 复合物能把QH 2 分子上的两个氢原子上的电子拿走,把这样形成的两个氢离子释放到细胞膜的外侧,再加上内侧形成QH 2 时消耗的两个氢离子,就相当于把两个氢离子从细胞膜的内侧转移到外侧了。
QH 2 是具有还原能力的分子,里面的能量还可以利用来产生跨膜氢离子梯度。这就是 bc 1 复合物的功能,和本章第九节图2-22里面的 bc 1 复合物非常相似。这个复合物由多个蛋白亚基组成,其中能够传递电子的蛋白质亚基都含有辅基,例如含血红素的细胞色素 b 和细胞色素 c 1 。另一个电子转移蛋白含有由铁原子和硫原子组成的电子传递辅基,叫做铁硫蛋白(Fe-S)。电子经过这些电子转移中心时,释放出来的能量被用来把氢离子从细胞膜的内侧泵到外侧,进一步增加细胞膜外侧氢离子的数量。释放了能量的电子被传递到复合物外一个能够在膜的外表面自由移动的蛋白质叫细胞色素 c 2 ,细胞色素 c 2 再把电子传回光反应中心失去了电子的叶绿素分子上,完成一轮电子循环。得到了电子的叶绿素恢复中性,又可以被光激发,开始另一轮电子循环。
这个系统虽然可以工作,但是效率不高。因为叶绿素分子太小,卟啉环的面积只有1平方纳米左右,这么小的面积接收不到许多光子。为了增加吸收光的面积,原核生物还发展出了捕光复合物。捕光复合物也由叶绿素结合蛋白(chlorophyll binding protein)组成,上面结合了12个叶绿素分子和两个类胡萝卜素分子。它们形成特殊的空间排列,以便使任何受到光子激发的分子都可以把能量传给光反应中心,在那里激发出电子。这相当于收集太阳光的“天线”,能够大幅度地提高光反应中心的工作速度。这种能量传递的效率非常高,在不到1纳秒的时间内就能把收集到的能量传输给反应中心,传输效率为95%,即基本上没有能量损失。有了捕光复合物,光反应中心的工作效率就可以大大提高了。所以原核生物最简单的光合反应系统是由捕光复合物、光反应中心、 bc 1 复合物组成的,工作方式是由光激发的电子循环。
这种光合反应系统是比较原始的,只含有一种光反应中心。它能够将光能转换成为跨膜氢离子梯度,效果和使用视紫红质的嗜盐菌相同,不能提供有机合成所需要的氢原子,也不释放出氧气,为一些细菌如紫细菌所使用。但是它创造的光反应中心和捕光复合物,却是伟大的发明,奠定了蓝细菌和植物中更复杂的,能够同时提供能量和氢原子的光合反应系统的基础。
紫细菌的光反应中心虽然可以产生QH 2 ,但是其中的氢原子并不被用于有机合成,而是像在呼吸链中一样,被用来产生跨膜氢离子梯度,有机合成所需要的氢原子仍然需要从现成的有机物来,例如葡萄糖、乙酸、琥珀酸,所以这样的细菌还摆脱不了消费者的角色。而如果世界上的生产者不多,消费者也不会繁荣,所以地球上还需要能够不消耗现成的有机物,而是能够自己用无机物合成有机物的生物。
为有机合成直接提供氢原子的主要是两种彼此类似的分子,烟酰胺腺嘌呤二核苷酸(Nicotinamide adenine dinucleotide,NAD + )和烟酰胺腺嘌呤二核苷酸磷酸(Nicotinamide adenine dinucleotide phosphate,NADP + ),后者比前者在核糖上多一个磷酸根。读者不必记住它们冗长复杂的名称,只需记住它们常用的名字,NAD + 和NADP + 就行了。它们都不是什么全新的分子,而是在ADP分子上再加上与核糖相连的烟酰胺,所以也是与核苷酸有关的分子。它们烟酰胺的部分可以反复地接受两个电子和一个氢离子,形成它们的还原形式——NADH和NADPH,它们就能够为有机合成提供氢原子(图2-32)。NADH和NADPH之间还可以互相转化,即NADH上的氢原子可以转移到NADP + 上,使其变为NADPH;NADPH也可以使NAD + 变成NADH。所以一旦有一种形式出现,只要细胞里有催化它们之间相互转化的酶,就可以生成另一种形式。在“消费者”生物中,它们是接受有机分子的代谢中产生的氢原子而形成的(例如参看图2-23三羧酸循环)。如果不依靠现成的有机物,能够从光反应系统直接生成NADH或者NADPH,生物就可以只依靠光能和无机的碳源(例如二氧化碳)和氮源(例如氮气和氨)等来合成有机物。这就是真正的有机物的生产者。
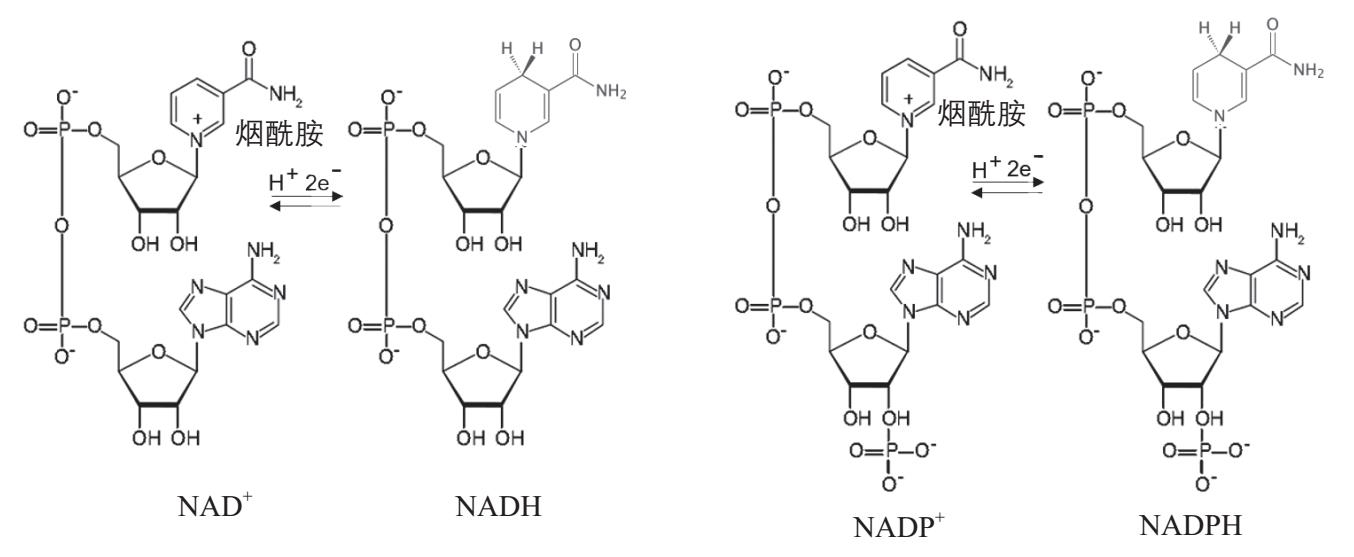
图2-32 NAD + 和NADP + 。它们分子下半部分都含有ADP,上半部分是烟酰胺通过核糖与ADP部分相连,NADP + 在核糖的2位羟基上还连一个磷酸根。它们烟酰胺部分都含一个正电荷。在接收到两个电子和一个氢离子后,它们就分别变为还原型的NADH和NADPH。NADH和NADPH都能够为有机合成提供氢原子
这样的光反应系统还真的出现了。这就是对原有的光反应中心加以改造,让叶绿素受激发时射出的电子不是以醌分子为受体,而是经过铁氧还蛋白还原NADP + ,形成NADPH。这个发明看来是在蓝细菌中完成的,因为蓝细菌就含有两种光反应中心,一种以醌为电子受体,一种以铁氧还蛋白为电子受体(图2-33)。
随着第二个光反应系统的出现,蓝细菌的细胞还发生了另一个变化,就是原来含有光反应系统的细胞膜向细胞内伸出,与其余的细胞膜分离,成为细胞内的膜。这些膜将一部分细胞质包裹起来,与其余的细胞质分开,叫做“类囊体膜”(thylakoid membrain)。由于光反应系统不再位于细胞膜上,光反应泵出的氢离子也不再被泵到细胞外,而是到类囊体膜围成的腔内。氢离子从类囊体腔流回细胞质时,带动ATP合成酶合成ATP。虽然紫细菌把氢离子从细胞内泵到细胞外,蓝细菌把氢离子从细胞质泵到类囊体腔,但都是把氢离子泵离细胞质。这个变化也为后来进行光合作用的真核细胞继承,成为叶绿体中的类囊体(参看图3-23)。
由于以铁氧还蛋白为电子受体的光反应中心被发现在先,被称为光系统Ⅰ(photosystem Ⅰ,简称PSⅠ)。以醌为电子受体的光反应中心本来是首先出现的,在结构和功能上都类似于紫细菌的单光反应中心,但是因为被发现在PSⅠ之后,被称为光反应中心Ⅱ(PSⅡ)。不仅如此,PSⅠ中的叶绿素被光激发射出电子后,产生的正电荷是被来自PSⅡ的电子中和的,其间经过 bf 复合物(相当于紫细菌的 bc 1 复合物)和一个叫做质体蓝素(plastocyanin,简称pc)的蛋白质,这样这两个光反应中心就被串联在一起,协调工作,而不是彼此单独活动。蓝细菌的双光反应中心系统既能形成跨膜氢离子梯度来合成ATP,又能提供用无机物合成有机物所需要的氢原子,使得真正的利用光能的有机物生产者出现,对地球上生物的发展与扩张具有关键性的意义。
不过把两个光反应中心串联起来也会产生一个问题,就是PSⅡ射出的电子会通过质体蓝素PC传给PSⅠ,不能通过环状电子流动流回PSⅡ中失去电子的叶绿素分子上,这些正电荷就不能完全被中和。在这里蓝细菌又完成了另一个伟大的发明,就是让PSⅡ中失去电子的叶绿素分子从水分子中的氢原子那里得到电子,把氢原子变成氢离子,释放到细胞膜外。失去了氢原子的氧原子彼此结合,以氧气的形式放出。这就是释氧光合作用(oxygenic photosynthesis)。释氧光合作用的出现是生物演化史上的重大事件,从此地球的大气中就有了源源不断的氧气供给。这就为本章第九节中说的“氧化磷酸化”提供了条件和保证。由于氧化磷酸化比底物水平磷酸化能够生成多得多的ATP,这就使得生物的进一步演化成为可能。而这个功绩也是由原核生物,特别是蓝细菌完成的。真核的藻类生物和植物的光合作用系统,就是从蓝细菌那里继承下来的,并且将其发扬光大,造成了地球上生物的大繁荣。