
下载掌阅APP,畅读海量书库
立即打开



原核生物的细胞里面含有DNA、RNA和蛋白质等生物大分子,以及一些盐类分子如氯化钠、氯化钾等。原核生物要生存,一个必要条件就是细胞的内容物要和环境分开,这样细胞里面的分子才不会被稀释到环境中去。这就需要原核生物有一个自己的“墙壁”,由它包围形成一个属于生物自己的小空间,这就是细胞。出于这个原因,所有的原核生物都是由细胞构成的。另一方面,原核生物也需要细胞外的物质源源不断地进入细胞,新陈代谢产生的废物也需要不断地被排出到细胞外面。这就要求这个“墙壁”不能“密不透风”,而是有“门户”供物质进出。在第一章第三节中我们已经谈到,模仿星际空间的条件,就可以生成能够在水中形成小囊泡的物质(见图1-8),说明组成细胞膜的物质可以在自然条件下生成。原核生物又将这些物质完善,完美地解决了细胞膜的“墙壁”作用和通透性的问题,这就是由磷脂组成的细胞膜和它上面的通道蛋白质。
我们在第一章第三节里讲过,亲脂力(弥散的电荷作用力)和亲水力(相对固定的点电荷之间的作用力)彼此配合,就有可能在水中形成空间结构。如果在一个分子上既有亲水的部分,又有亲脂的部分,即两性分子,其亲水的部分可以处在结构表面,和水直接打交道,而亲脂的部分由于受到水分子的排斥,被“赶”到一起,处于结构内部,彼此以色散力相吸引,并且从内部“拉住”分子的各个部分,这两种作用相互配合,就能在水中形成相对稳定的结构。
脂肪酸(fatty acid)是两性分子。它的亲脂部分是由16或18个碳原子组成的长链,上面再连上氢原子。它的亲水部分是一个叫做“羧基”的“头部”(见图1-7)。我们可以把这个分子想象成一根火柴,火柴杆亲脂,火柴头亲水。在中性(pH 7)的环境中,羧基上的氢原子会有一部分把电子完全给氧原子,形成氢离子而脱离脂肪酸,这样失去氢离子的脂肪酸头部就会带负电。当脂肪酸的分子被放到水中时,亲脂的火柴棍被水“排挤”,彼此聚到一起,通过色散力相互吸引,形成一个亲脂的内部。而亲水的火柴头则排列在外面,与水亲密接触。对于亲脂的火柴杆来讲,它们越是整齐排列,色散力越强,所以火柴杆趋向于彼此平行。而火柴头由于带负电,彼此排斥,趋向于增加火柴杆之间的距离。这两种力量平衡的结果就是形成一种球形结构,火柴杆指向球心,火柴头排列在球面上。脂肪酸在水中也可以形成双层膜,包裹一些水在内部,形成小囊泡(见图1-7)。不过由脂肪酸形成的囊泡不很稳定,内部空腔很小甚至没有空腔,作为细胞膜不是很理想的,原核生物的细胞膜也不是由脂肪酸组成的。
如果亲脂的部分增加到两根火柴杆,而且共用一个火柴头,火柴杆之间的吸引力就会更强,更容易彼此平行排列,而火柴头占的空间是原来的一半,彼此的排斥力也会更弱。这两种力量变化的结果就是更容易形成近似平面的结构。但是火柴杆的另一头由于没有亲水的头部,仍然要与水接触,而这种情况是不稳定的。一个解决办法是平面结构中火柴杆没有头的那一端与另一个平面结构的火柴杆“足和足”相对,彼此接触。这样两片膜就对在一起,每层膜亲脂的火柴杆朝内,彼此接触;亲水的火柴头都向外,面对水,把内部的脂层和外面的水隔开,形成双层膜,这样就解决了亲脂层与水接触的问题。不过这样的膜还有一个麻烦,就是在膜的边缘,亲脂的部分仍然会暴露出来,和水接触。为了避免这种情况,膜会由于热运动而自动弯曲,让边缘融合,形成小囊泡,这样亲脂的部分就完全和水隔绝了。原核生物的细胞膜就是这样形成的。这样形成的囊泡尺寸较大,可以满足细胞空间的要求。
能够使两个脂肪酸尾巴共用一个亲水“头”的分子就是磷脂(phospholipid)。磷脂分子的核心是一个甘油分子。它有三个碳原子,每个碳原子上面连上一个羟基,其中两个羟基(包括位于中间位置上的羟基)分别和两个脂肪酸分子的羧基作用,形成“酯键”,把两个脂肪酸分子连在甘油分子上。第三个羟基则和一个磷酸分子相连,磷酸再和一个亲水的分子(如胆碱)相连。这样形成的分子就叫磷脂(图2-16)。
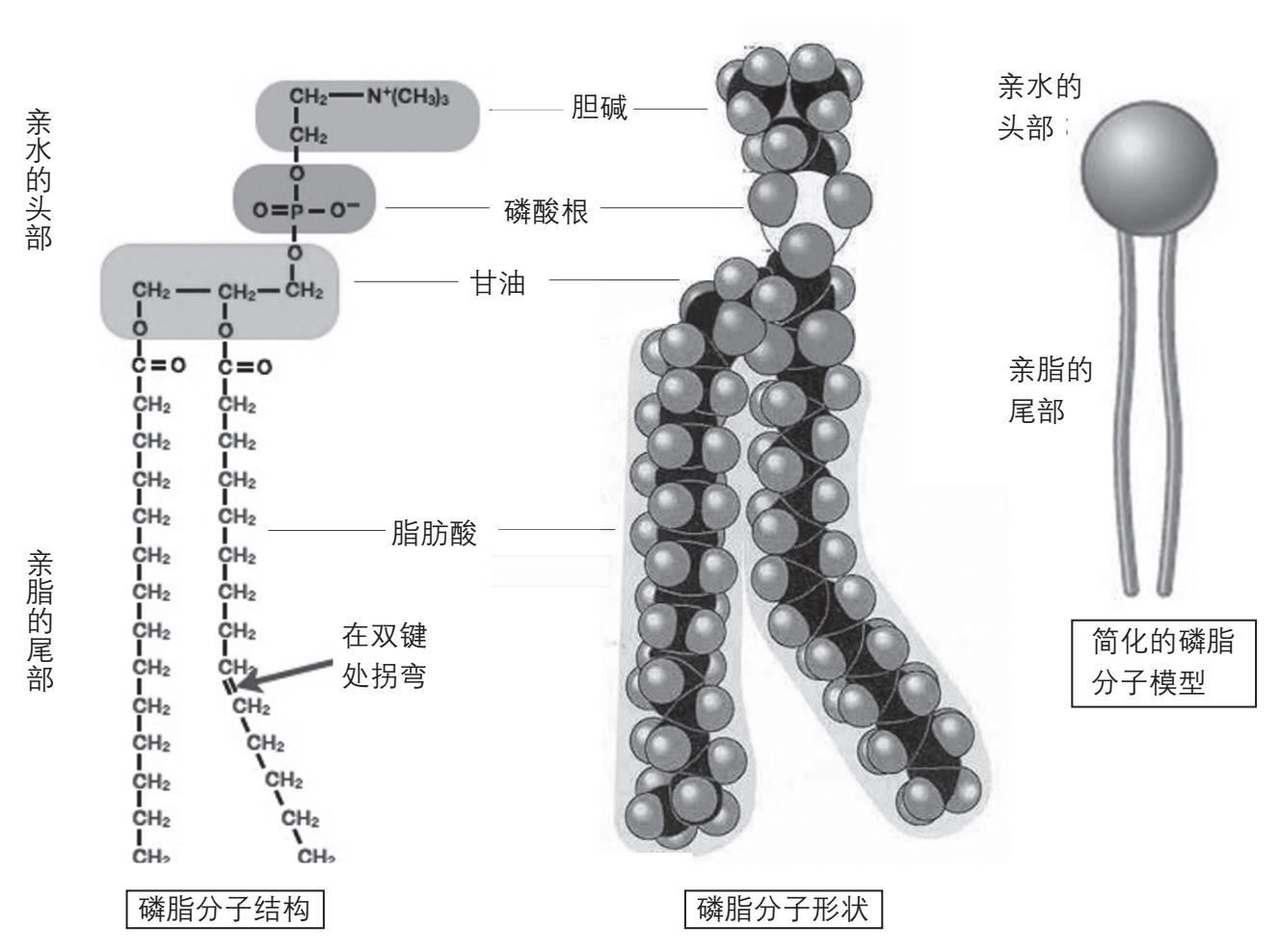
图2-16 磷脂分子。磷脂分子由甘油分子连上两个脂肪酸分子和一个磷酸分子,磷酸分子再连上一个亲水分子组成。左图为分子结构图。中图为分子模型,注意脂肪酸中间的一个碳-碳双键使脂肪酸分子有一个“拐弯”。右图为简化的磷脂分子模型
就像前面说的那样,磷脂有两个脂肪酸“火柴杆”和一个亲水的头部,它在水中就可以形成双层膜。双层膜的总厚度约5纳米(5×10 -9 米),其中亲脂的内层厚2.5纳米,相当于25个氢原子的大小。这层膜就能够防止DNA、RNA、蛋白质这样的生物大分子“逃”到细胞外面去,相当于细胞的“墙壁”(图2-17左)。
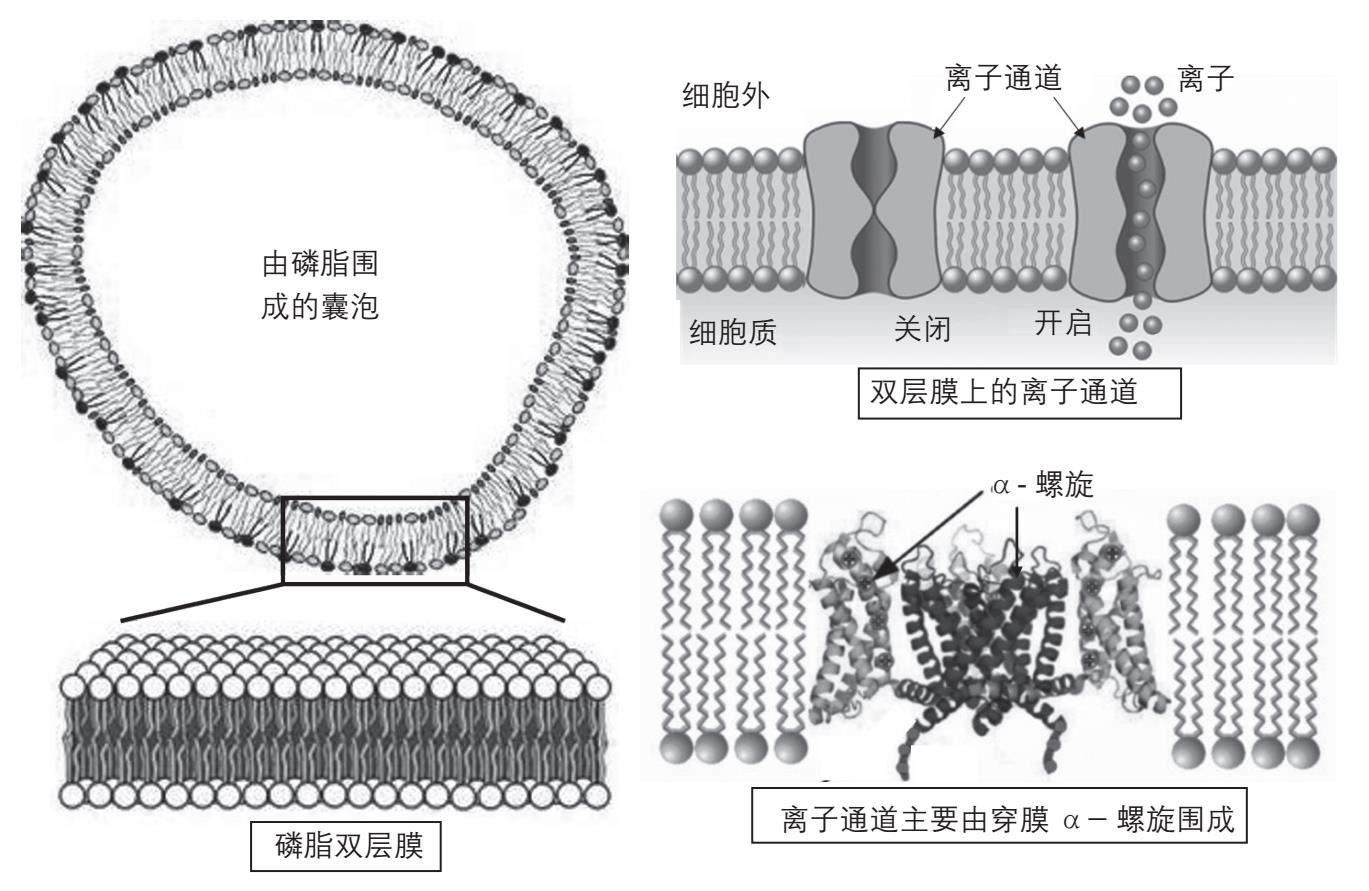
图2-17 由磷脂分子组成的双层膜和小囊泡以及膜上的离子通道。左上为磷脂双层膜围成的囊泡,可以起到细胞膜的作用,左下为磷脂双层膜的结构图。右上:磷脂膜上有各种通道让物质进出细胞,包括离子通道。这些通道可以根据不同的情况关闭或者开启。右下:这些离子通道主要由穿膜的 α-螺旋围成
“墙壁”的功能解决了,那么“门户”的功能呢?有些分子是可以通过扩散直接穿越细胞膜的,并不需要“门户”,比如氧分子和二氧化碳分子。但是许多分子,特别是高度水溶性的分子如葡萄糖,以及带电的离子如钾离子和钠离子,就要靠镶嵌在细胞膜上的蛋白质分子进出细胞。这些蛋白质分子贯穿细胞膜,沟通细胞内外。细胞需要的分子和离子可以通过这样的蛋白质通道进入细胞。这些镶嵌在细胞膜上的蛋白质和溶解于水中的蛋白质不同,叫做膜蛋白(membrane protein)。在这里蛋白质遇到了不同的环境:即有25个氢原子厚的“油层”。为了穿过这些“油层”,蛋白质分子有一个或多个区段,里面的侧链多数是亲脂的。这些亲脂节段形成亲脂的α-螺旋,可以容易地“穿过”细胞膜,而蛋白质中其余带有许多亲水侧链的节段则位于细胞膜之外。当一个膜蛋白有多个“穿膜节段”时,这些“穿膜节段”也含有少数亲水的侧链。这些亲水侧链在脂性环境中被排斥,彼此通过固定电荷相互吸引,使这些穿膜节段彼此靠近,围成管状,形成离子通道(ion channels)(图2-17右)。在这里蛋白质穿膜节段中亲水和亲脂侧链的位置就反过来了:亲脂的侧链朝外,与膜的脂性环境接触;亲水的侧链朝内,形成离子通道。所以膜蛋白的结构也是由亲水和亲脂这两种作用力相互配合形成的,不过由于环境不同,穿膜节段的朝向和水溶性蛋白正好相反。这些通道还可以根据需要被打开和关闭,就像房间的门窗可以根据需要开关一样。这样,“墙壁”和“门窗”的功能都具备了。
如果检查组成细菌细胞膜的磷脂里面的主要脂肪酸分子,发现它们都很长。比如棕榈酸和软脂酸有16个碳原子,油酸、亚油酸、亚麻酸和硬脂酸都有18个碳原子。从原核生物中的细菌到真核生物再到人,磷脂里面的主要亲脂部分都是由这些16或18碳的脂肪酸组成的。这些长链脂肪酸都是高度不溶于水的,合成、吸收和运输都很麻烦,为什么生物要用这么长的脂肪酸呢?
主要原因有两个。一是细胞膜必须足够“结实”。细胞膜是细胞对外的“屏障”,容不得出任何差错。细胞膜破裂往往意味着细胞死亡。双层膜只由两层分子组成,是很薄的,比肥皂泡的膜(大约700纳米厚)还要薄。如果把细胞放大成为直径4米的房间,细胞膜的厚度只相当于1毫米!这么薄的细胞膜除了要经受周围分子的热运动造成的冲击,还要耐受细胞内容物造成的渗透压,只有16到18碳原子长的脂肪酸才可以产生足够强的色散力,使碳氢链“火柴杆”之间的作用力足够强,使细胞膜足够强固。17碳以上的烷烃(饱和的碳氢链),在常温常压下已经是固体。为了不让细胞膜真的成为“固体”,细胞膜已经采取了多种措施来保持其流动性,包括使用不饱和脂肪酸(含有碳-碳双键的脂肪酸,双键会在“火柴杆”上引起“拐弯”,见图2-16和图1-2)来扰乱脂肪层的结构。这意味着原核细胞已经把脂肪酸的长度推到形成“固体”的边缘,以求得足够的强度。
第二个原因是细胞膜必须成为离子的有效屏障。细胞内外的离子种类和数量的差别是很大的。比如大肠杆菌和人体的细胞一样,细胞内有高浓度的钾离子和低浓度的钠离子,细胞外相反,有高浓度的钠离子和低浓度的钾离子。这种膜两边离子浓度的差别对细胞的生理功能极为重要,所以细胞膜必须要能够防止离子“泄漏”。25个氢原子厚的脂质层对离子来讲就是脂肪的“汪洋大海”。即使是这样,轻度的“泄漏”仍在发生,要靠“离子泵”不断地把泄漏的离子“泵”回去。要是膜再薄,膜两边离子的浓度差就难以维持了,细胞也会因为要消耗太多的能量来维持膜两边离子的浓度差而“累死”。
迄今为止,我们只谈到原核生物中的细菌的细胞膜,而没有谈到同为原核生物的古菌的细胞膜。这是因为古菌细胞膜的构造比较特别,不仅与细菌的细胞膜不同,与真核生物的细胞膜也不同。大概是由于古菌常常生活在极端严酷的环境中,例如高温、高压、高盐浓度,高酸性环境或高碱性环境等,在这些不利条件下磷脂中脂肪酸和甘油之间的酯键容易被水解,使细胞膜的通透性增加,甚至造成细胞膜破裂。为了适应这些不利的环境,古菌不使用脂肪酸来构建磷脂,而是用聚异戊二烯的长链通过醚键(—C—O—C—)和甘油相连。这样的分子也能在水中形成双层膜结构,但是由于醚键比酯键稳定得多,亲脂的“火柴棍”还带侧链,使得古菌的细胞膜更加结实稳定。古菌也含有使脂肪酸降解的酶,说明脂肪酸也是古菌代谢系统中的一部分。古菌不用脂肪酸来建造细胞膜,也许是古菌在适应严酷环境的过程中放弃了使用含脂肪酸的磷脂,而采用聚异戊二烯的脂肪链(图2-18)。
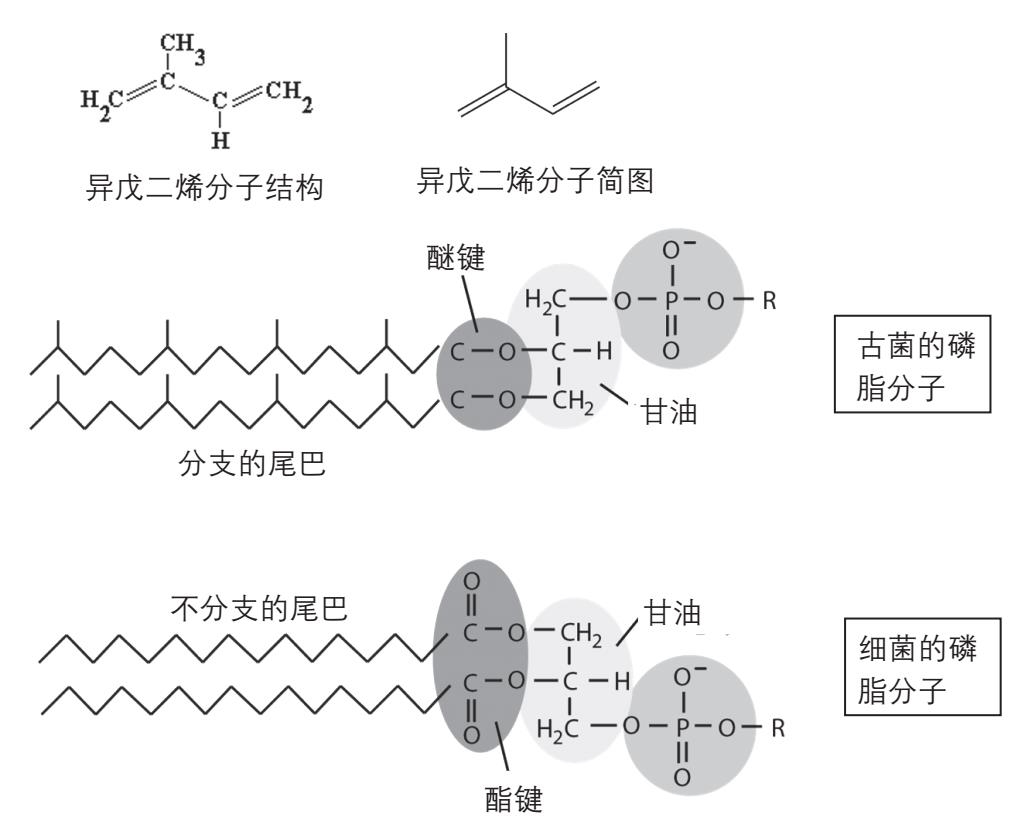
图2-18 古菌的磷脂分子。与细菌的磷脂分子(下)相比,聚异戊二烯的分支链取代了脂肪酸的直链,醚键代替了酯键。异戊二烯聚合时,双键会消失,产生分支的饱和碳氢链
含有膜蛋白的磷脂双层膜是原核生物对生命发展的又一重大贡献。真核生物,包括植物、动物和人,都几乎原封不动地继承了细菌细胞膜的这种构造,连使用的主要脂肪酸(硬脂酸、软脂酸、油酸、亚油酸)都相同。许多细菌为了更好地保护自己,在细胞膜的外面还有其他的保护层,例如荚膜和细胞壁。植物细胞为了增加机械强度,外面也有细胞壁(cell wall)。虽然它们的名字叫做“细胞壁”,这些细胞膜外面的结构却只有保护和增强的作用,并不能选择性地阻挡和允许分子和离子通过,所以不是细胞真正的“墙壁”和“门户”。动物的细胞则只有细胞膜,除少数细胞如神经细胞外,细胞膜外也没有更多的包裹层,这说明细胞膜才是细胞最基本的屏障和与外界交换物质的通道。