
下载掌阅APP,畅读海量书库
立即打开



 T淋巴细胞、B淋巴细胞与免疫应答
T淋巴细胞、B淋巴细胞与免疫应答
拉夫
编者按
在这篇文章中,出生于加拿大的细胞生物学家马丁·拉夫就T淋巴细胞和B淋巴细胞已有的知识进行了总结。在外周淋巴器官中发现了两种不同类型的白细胞,这两类白细胞在外周淋巴器官中看起来是一样的。拉夫在之前的研究中就已经鉴定了一种T细胞标志物,并因此立即获得国际认可。T细胞和B细胞具有不同的起源、特性和免疫功能,两者在功能上是相互调控的。拉夫说他们的这一发现标志着免疫学的新时代。在这个新时代中,强大的研究工具和可利用的研究模型很有可能为生物学问题和疾病的研究带来曙光。 英文

认识到两类截然不同的淋巴细胞是免疫学研究中一个转折点。免疫学模型和工具将有助于解答多种生物学问题。 英文

虽然免疫学已经发展成了一门独立的振奋人心的学科,但是关于淋巴细胞及其所介导的免疫应答的研究对医学和生物学其他分支学科也具有重要的影响。然而遗憾的是,免疫学独有的语言体系使得非免疫学家很难参与到这个振奋人心的领域中。因此,我们撰写了这篇综述文章,力图对已知的免疫学的细胞学基础进行概括性阐述。(关于淋巴细胞和它们在免疫应答中的作用,更详细的综述见参考文献1。) 英文
免疫学是一门研究动物对进入其体内的外源性物质(抗原或免疫原)所产生的特异性反应的科学。所有的脊椎动物都可以产生这种免疫应答。这个过程包括特异性免疫球蛋白分子(抗体)和(或)具有特异性反应活性的细胞的产生。它们都可以在血液中循环,并与抗原特异地结合。这种反应会使外源性物质失活(如细菌毒素)、被杀死(如受感染的器官或移植的细胞)和(或)被网状内皮系统中的细胞吞噬。另一方面,在某些情况下,这种免疫应答可能会对宿主自身产生有害的作用。例如在超敏反应中(比如花粉过敏和药物过敏),抗原与吸附在嗜碱性粒细胞和肥大细胞上的抗体发生反应,从而引起组织胺和其他药理炎症介质的释放。一般来讲,那些可以通过将致敏供体(携带抗体)的血清转移到另一个动物体而引起的免疫应答称为体液免疫(或抗体)反应,而那些不能通过血清只能通过致敏细胞才能转移的免疫应答则被称为细胞免疫反应。 英文
二十世纪五十年代和六十年代早期,当免疫化学家们正在努力解析抗体结构的时候,细胞免疫学家们正致力于阐明淋巴细胞是参与免疫反应的主要细胞。最具说服力的实验是在大鼠中进行的:研究人员首先利用放射性射线照射杀死了受体大鼠的淋巴细胞从而使其自身无法进行免疫应答(见参考文献2),然后将从大鼠的主淋巴管—胸导管中分离出来的相对纯的淋巴细胞移植到受体大鼠体内,结果发现这样可以使受体大鼠同时获得细胞免疫和体液免疫。此外,研究人员还发现通过胸导管持续引流使动物体内的淋巴细胞耗尽会削弱免疫反应性 [2] 。至此,长久以来起源和功能一直是个谜的淋巴细胞,终于被确认为“免疫活性”的细胞。 英文
很快人们便认识到,淋巴细胞并不是一个同质的群体。许多证据表明存在两种截然不同的具有免疫活性的淋巴细胞:一种淋巴细胞需在胸腺中发育并与细胞免疫有关;另一种淋巴细胞的发育则与胸腺无关并且介导体液免疫应答。支持这一观点的证据来自于二十世纪六十年代在鸟类、啮齿动物以及人类中的研究:在鸟类 [3,4] 和啮齿动物 [5] 中,摘除胚胎或新生动物的胸腺,它们长大后细胞免疫应答会显著受损,而对体液免疫影响不大。另一方面,在孵化阶段摘除法氏囊 [3,4] (鸟类特有的泄殖腔淋巴器官)会削弱鸟类产生抗体的能力,而几乎不影响细胞免疫。对免疫缺陷病患者的研究也表明,体液免疫和细胞免疫分别受到不同因素的影响(见参考文献6):由于缺乏产生抗体的淋巴细胞,患有布鲁顿型先天性丙种球蛋白缺乏症的病人不能产生抗体,但是细胞免疫正常;而先天性胸腺发育不全的儿童(如迪乔治综合征患者)则表现为细胞免疫功能显著受损,但对某些抗原产生免疫应答时可以产生相对正常量的抗体。 英文
在过去的几年中,免疫的双淋巴细胞模型(至少在鸟类和哺乳动物中)已经确立。在这个模型中,有两个“中枢”淋巴器官——法氏囊或在哺乳动物中具有的同功能器官(目前尚不明确)和胸腺——产生不依赖于抗原的淋巴细胞,并将它们运送到“外周”淋巴器官(淋巴结、脾脏和肠道相关淋巴组织),淋巴细胞在那里等候与抗原接触从而诱导其进一步分化为“效应”细胞(见下文)。其中,在外周淋巴组织中胸腺来源的淋巴细胞被称为T细胞,而鸟类的法氏囊或哺乳动物相应器官来源的淋巴细胞被称为B细胞 [7] 。 英文
直到最近,人们都一直认为只有脊椎动物才具有特异性免疫反应。然而,现在有证据表明,诸如环节动物和被囊动物等无脊椎动物也可以排斥外源组织,并且这种原始的免疫应答也具有一定的特异性和短期的记忆效应 [8] (即在第二次接触到相同抗原时可以产生更强和(或)更快的免疫应答)。这些反应是由巨噬细胞样细胞(体腔细胞)介导的,也可能是由特异性相对较差的可溶性效应分子介导的 [8] 。由于并没有证据表明无脊椎动物中具有淋巴细胞或免疫球蛋白,因此特异性细胞免疫很可能是在脊椎动物的两种主要免疫细胞出现之前形成的。 英文
所有脊椎动物都有淋巴细胞,也可能有胸腺组织(至少在发育的某个阶段中),并且可以产生抗体、引发细胞免疫应答 [8] 。低等脊椎动物(如七鳃鳗类和盲鳗类)几乎没有系统化的淋巴组织且只能产生一类抗体(IgM样抗体)。初级淋巴结样结构是在两栖动物中首次发现的,这种结构可以产生两类抗体。第一种被发现具有明显两类淋巴系统的脊椎动物是鸟类;而且鸟类独一无二地具有两个独立的中枢淋巴器官——胸腺和法氏囊,二者可以分别产生T细胞和B细胞。尽管B细胞的发育位点尚未确定,但可以肯定的是哺乳动物具有大量的、高度系统化的淋巴组织,可以产生各种不同种类的抗体(例如人体可以产生IgG、IgM、IgA、IgE、IgD等),并且具有截然不同的T细胞和B细胞群。我们还不知道比鸟类低等的脊椎动物是否具有独立的T细胞和B细胞群。 英文
在大多数动物中,淋巴细胞首先在胎儿胸腺中出现。胸腺原基由上皮细胞组成,来源于第三、第四咽囊。过去人们认为胸腺淋巴细胞(胸腺细胞)是由胸腺上皮细胞发育而来,但是鸡和小鼠的实验清楚表明,胎儿卵黄囊和肝脏中的造血干细胞会迁移到胸腺原基并在那里增殖、分化成胸腺淋巴细胞,这些过程大概是在胸腺上皮诱导作用下进行的 [9] 。在小鼠(妊娠20天)中,在胚胎期第11天左右,可以观察到首批类似大嗜碱性母细胞的干细胞到达胸腺;而在胚胎期第15天或第16天的时候可以检测到首批小淋巴细胞 [9] 。利用放射性示踪 [10] 、染色体 [5,11] 和细胞表面抗原标志物 [9] 可以清楚地显示淋巴细胞从胸腺迁移到外周淋巴组织并分化为T淋巴细胞群。尽管这一过程在小鼠临出生前开始,但是大部分迁移过程发生在其出生后的第一周 [9] 。因此,如果在小鼠出生的第一天便摘除其胸腺,那么其长大后则表现为明显的T细胞缺陷从而损伤细胞免疫;然而如果晚些时候进行胸腺切除,那么影响会小很多 [5] 。在成年动物中,骨髓来源的干细胞会迁移到胸腺,胸腺淋巴细胞也会持续地向外周组织迁移。但是与胚胎和新生动物相比,成年动物的这一过程要缓慢得多 [5,11] 。 英文
大多数胸腺淋巴细胞没有免疫活性(也就是说它们不能对抗原作出应答),并且在其他方面它们与外周T淋巴细胞也有所不同,这表明从胸腺细胞到T细胞存在另一个分化过程。最近有研究表明,在胸腺髓质中存在胸腺细胞小的亚群(约2%~5%),它们具有免疫活性并且具有外周T淋巴细胞的大部分特征 [9,12,13] 。这表明第二个分化步骤可能是在胸腺内完成的,可以把T细胞的发育看作如下过程:干细胞→胸腺细胞→“成熟”胸腺淋巴细胞→外周T淋巴细胞(见图1)。但是几乎可以肯定的是,上述模型只是一种过于简化的说明。因为有证据表明,细胞在成熟的各个阶段细胞都可能离开胸腺,或者作为不同的细胞系产生若干具有不同特性和功能的外周T细胞亚群 [13] 。另外,人们假定的胸腺体液因子或激素(例如胸腺素)的功能仍不清楚。尽管有证据表明在胸腺外它们很可能不能诱导干细胞分化为淋巴细胞,但它们可能以某种方式影响着外周T淋巴细胞 [14] 。 英文
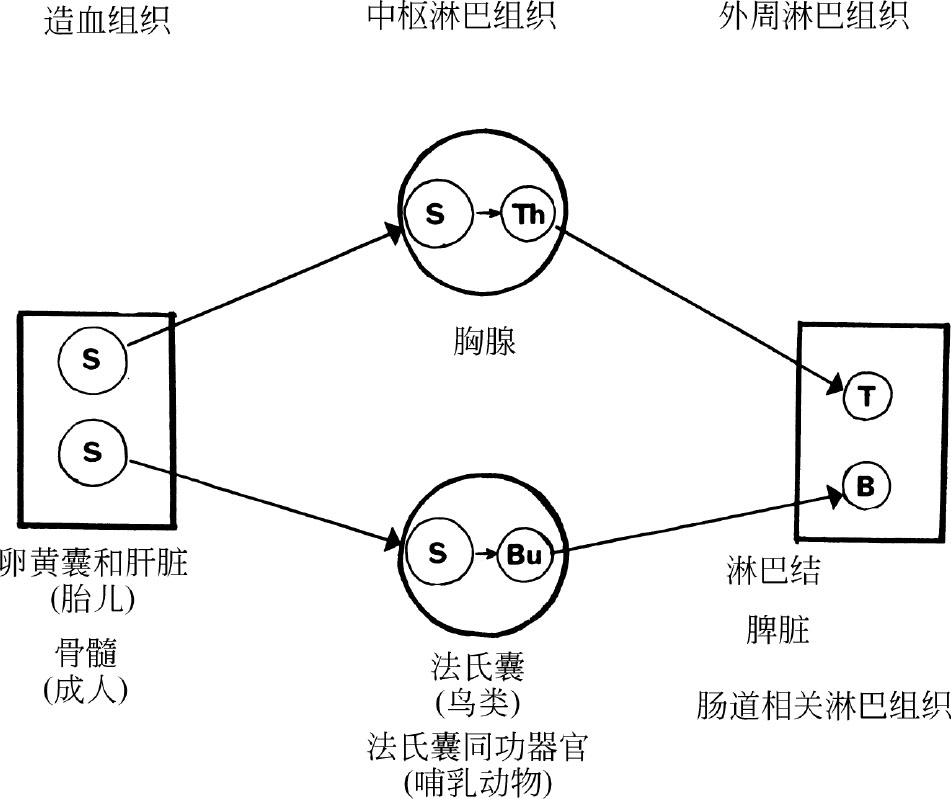
图1. T淋巴细胞和B淋巴细胞的发育过程示意图(过于简化的——见正文)。干细胞(S)迁移到胸腺和法氏囊组织,并在那里分别分化成胸腺淋巴细胞(Th)和法氏囊淋巴细胞(Bu)。然后这些细胞的一部分迁移到外周淋巴组织中,分别成为T淋巴细胞和B淋巴细胞。
在鸟类中,B细胞的发育依赖于法氏囊。法氏囊是在胚胎发育第五天由泄殖腔背壁外翻所形成的一个囊状结构。染色体标记研究显示,在胚胎发育第12天到第13天时便有干细胞(与胎儿胸腺中检出的干细胞形态一致)从卵黄囊迁移到法氏囊中,并且在1~2天内在那里分化成淋巴细胞 [9] 。到第14天可以观察到表面有IgM的法氏囊淋巴细胞,几天后可以观察到产生IgG的法氏囊淋巴细胞 [15] 。同位素标记实验证明法氏囊淋巴细胞可以迁移到外周淋巴组织。在胚胎时期摘除法氏囊会导致外周B淋巴细胞的显著缺失和抗体(即免疫球蛋白)产量的显著减少 [15] 。最近人们发现,向孵化前的胚胎注射抗μ抗体(特异靶向IgM重链)加上在胚胎孵化后摘除其法氏囊,抑制IgM的同时,也会抑制随后IgG的产生 [15] 。这表明即使是那些最终产生IgG的B细胞,最初在其表面也会表达IgM;同时强有力地证明,单个的B细胞中存在IgM到IgG的转换。但是这种转换是由抗原驱动(如小鼠实验结果显示 [16] ),还是与抗原刺激无关(如鸡实验结果显示 [15] ),目前尚不清楚。 英文
在哺乳动物中,人们尚不清楚干细胞是在哪里分化成B淋巴细胞的,只知道肯定不是在胸腺中。有人认为哺乳动物的肠道相关淋巴组织(如派伊尔节、扁桃腺、阑尾等)可能充当“法氏囊同功结构”,不过尚缺乏证据证明这一点。但是至少在啮齿类动物中,越来越多的证据表明大量的淋巴细胞是在造血组织中(如胚胎的肝脏、成年动物的骨髓)产生的 [17] ;并且似乎这些组织不仅为T细胞群和B细胞群提供干细胞,而且也是干细胞分化为B淋巴细胞的场所。 英文
目前还不清楚哪个阶段的干细胞会分化为淋巴细胞或成为T细胞、B细胞。研究人员在早期小鼠胚胎胸腺中观察到了多能造血干细胞(即可以分化为任何一种成熟血细胞、淋巴细胞或骨髓细胞的细胞) [18] ,这表明干细胞直到进入胸腺或法氏囊(或法氏囊同功结构)微环境以前都不能完成分化。 英文
由于处于静息状态的T细胞和B细胞在形态上无法区分,并且都分布于外周淋巴组织中,因此寻找区别、分离它们的方法对研究其各自的特性来说极其重要。就这一点来说,揭示它们之间重要的表面差异尤为有效。有些表面差异可以通过抗体识别出来 [19] 。例如θ同种异型抗原(由同种抗体识别,用一个种系的小鼠胸腺细胞免疫另一个种系的小鼠得到该抗体)存在于小鼠胸腺细胞和T细胞,而不存在于B淋巴细胞,因此其被证实为小鼠T细胞合适的表面标志物 [19] 。另一方面,已确实证明的细胞表面免疫球蛋白(Ig)(参考文献20、21)和“小鼠特异性B淋巴细胞抗原”(MBLA)(参考文献19)可以作为B细胞的标志物,MBLA是通过兔抗鼠B细胞产生的异种抗体识别的异种抗原。在补体存在下,抗血清能与一种或其他类型淋巴细胞表面发生特异反应,杀死相应细胞群,从而将它们从细胞悬液中清除。利用耦联到可消化的固相免疫吸附剂上的抗体 [22] ,或荧光标记的抗体,通过荧光激发的电子细胞分选技术 [23] ,就可纯化出任意一种细胞类型。除了上述T细胞与B细胞的表面抗原差异外,B细胞还可以通过细胞表面的补体受体与抗体-抗原-补体复合物结合 [24] ,或通过复杂的免疫球蛋白的Fc片段受体与抗原-抗体复合物结合 [25] ;而静息状态的T细胞则不具有这些受体。虽然目前还不清楚B细胞上Fc片段和补体受体的功能,但有人认为它们可能在淋巴组织中抗原的定位、抗原激活B细胞和(或)一般认定的B细胞杀死抗体包被的靶细胞的过程中起重要作用。 英文
大多数T淋巴细胞都会在血液和淋巴之间反复循环。T细胞可以通过淋巴结和派伊尔节内特化的毛细血管后微静脉从血液中出来,穿过淋巴组织基质进入输出淋巴管;然后通过胸导管重新回到血液循环系统中 [2,5] 。尽管大多数B淋巴细胞似乎并不参与上述循环过程,但其中有些B淋巴细胞确实会通过淋巴组织的不同部位,以比T细胞慢的运送速度进行再循环 [26] 。在外周淋巴组织中,T细胞和B细胞的分布区域基本上是相互分隔的,分别为所谓的胸腺依赖区(脾脏动脉周围鞘、淋巴结副皮质区、胃肠淋巴组织的滤泡间区)和非胸腺依赖区(淋巴滤泡和脾脏白髓的外周区域、淋巴结中的滤泡和髓质以及胃肠淋巴组织滤泡) [27] 。将放射性标记的T细胞或B细胞注入动物体后,它们会特异地迁移到各自的区域 [27] 。尽管T淋巴细胞群和B淋巴细胞群都是异质的 [1] ,但是与B细胞相比,T细胞的平均增代时间更长 [28] ,并且体积略大 [29] ,密度更高 [24] ,粘附性更小 [24] (与玻璃、塑料、尼龙等各种材料的粘附性相比),带的负电荷也较多 [30] 。此外,T细胞更容易被抗淋巴细胞的血清清除 [31] (主要针对反复循环的细胞),但通常对细胞毒性药物(如环磷酰胺 [32] )、皮质类固醇 [33] 和辐射 [34] 的敏感性较低。另外它们在体外对于各种“有丝分裂原”的反应也是不同的,比如植物提取物(如植物有丝分裂原)、细菌代谢产物(如内毒素)或淋巴细胞表面抗原的抗体,其均可刺激很大一部分T和(或)B淋巴细胞分裂、分化为母细胞。尽管美洲商陆可以同时刺激T细胞和B细胞增殖,但是伴刀豆球蛋白A(Con A)、植物凝集素(PHA)和扁豆素只能刺激T细胞增殖,而脂多糖(如大肠杆菌内毒素)和抗免疫球蛋白血清则只能刺激B细胞增殖 [35] 。有意思的是,尽管可溶的ConA和PHA只能选择性地激活T细胞,但它们同样可以很好地与B细胞结合;而一旦将它们共价耦联到固相支持物上,它们也可以刺激B细胞增殖 [35] 。作为淋巴细胞被特异抗原活化的一种可能的模型,有丝分裂原刺激淋巴细胞活化被广泛研究。这些研究使得人们逐渐认识到淋巴细胞的激活不仅仅是配体与细胞表面受体结合那么简单。 英文
免疫学的中心法则是克隆选择学说。该学说提出在个体发生的一定时间内,单个淋巴细胞(或淋巴细胞克隆)通过细胞表面抗原特异性受体只对某一种或相对少数的几种抗原起反应,这一过程与抗原无关。于是,一旦抗原进入体内便会筛选出表面已有相应受体的淋巴细胞;抗原与受体的相互作用可以激活这些特异性细胞。现在关于T淋巴细胞和B淋巴细胞的克隆选择学说都有了令人信服的证据。因此,人们发现T细胞和B细胞都可以结合抗原至其表面 [36] (尽管曾经证明T细胞结合抗原比证明B细胞结合抗原要困难得多),并且通常只有一小部分的淋巴细胞(在未免疫的动物中约为万分之一到十万分之一)能够与任何抗原结合。此外,如果用强放射性抗原作用于淋巴细胞,无论是T细胞反应还是B细胞反应都会被选择性摧毁,而针对其他抗原的免疫反应却不会受到影响 [37] 。同样,如果把某种特定的抗原耦联到玻璃微珠上,那么可以与这种抗原反应的B细胞会特异性地黏附于玻璃微珠上,从而将它们从细胞悬液中特异地分离出来 [38] 。尽管出于某种未知原因,在此条件下T细胞通常不能黏附 [38] ,但用载有特异性同种抗原的单层细胞,可以选择性去除识别这种抗原的T细胞 [39] 。 英文
早在1900年,埃尔利希便提出那些可以产生抗毒素的细胞(现在知道是B细胞)在其表面有作为受体的抗毒素分子。这种受体学说的最新说法提出,B淋巴细胞表面有作为抗原受体的抗体分子(即免疫球蛋白),这些受体与B细胞或其子细胞最终分泌的抗体至少在结合位点上是相同的。现在有很好的证据支持这一观点:B细胞表面确实有免疫球蛋白分子(每个细胞大约10 4 ~10 5 个)(参考文献20、40),并且这些抗Ig的抗体抑制它们与抗原结合或应答的能力(有关综述见参考文献1)。此外,越来越多的证据表明,对于任何一个B淋巴细胞克隆来说,受体和分泌的抗体具有相同的抗原结合特异性 [41,42] 。但是,受体与最终分泌的抗体在免疫球蛋白亚型方面可能不总是一样的,比如一些分泌IgG细胞的B细胞前体似乎有IgM受体 [15,16] 。这些不同亚型的抗体(例如IgG与IgM)可能具有相同的抗原识别特异性(也就是说不同的免疫球蛋白恒定区可以与同一可变区组合 [43] ),而且单个克隆内IgM到IgG的转换并不意味着特异性的转变。至少在小鼠中有证据表明,大多数未经过抗原激活的B细胞表面都有IgM受体(以7-8S的单体形式存在 [44] ),而在与抗原初次接触后,这些B细胞则会转换类型 [16] 。关于抗体多样性是如何产生这一最根本的问题,也就是动物如何产生如此庞大的各种各样的免疫球蛋白分子(包括受体和分泌的抗体),目前仍存有争议。胚系学说认为动物生来就具有大量免疫球蛋白可变区基因;而体细胞突变理论认为动物生来只有很少的免疫球蛋白可变区基因,在随后的一些体细胞过程(例如突变或重组)中产生了大量可变区基因。 英文
T细胞表面受体的化学本质大概是当前细胞免疫学中最富争议性的问题。最简单也最合理的观点是:只有抗体可以识别抗原,而且所有抗原特异性受体必然是免疫球蛋白。但是由于许多研究人员都无法证明T细胞表面有免疫球蛋白的存在,或使用抗免疫球蛋白血清能抑制T细胞与抗原的作用,因此这一观点受到了质疑。事实上,越来越多的证据表明,至少在T细胞识别和(或)应答某些抗原中,除了传统的免疫球蛋白外,细胞表面组分也可能发挥重要作用。这些T细胞“受体”的优先候选分子很可能是免疫应答(Ir)基因编码的产物,Ir基因在遗传上与主要组织相容性基因座相关联 [45] 。这些Ir基因会影响T细胞对多种抗原的应答 [46] 。T细胞应答的精准特异性非常类似于抗体的特异性和B细胞识别的特异性 [47] ;再加上种种(尽管存有争议)实证表明T细胞上有免疫球蛋白(有关综述见参考文献1),使得人们不愿放弃T细胞表面具有免疫球蛋白受体的观点。也许T细胞(可能也包括B细胞)表面至少具有两套“识别”系统,其中一套包含免疫球蛋白,另一套通过Ir基因编码的产物介导,对于不同的抗原、不同的免疫应答和(或)不同的T细胞亚型来说,这两套识别系统可能具有不同的重要性。此外,这种假定的非免疫球蛋白识别系统可能与无脊椎动物对外源性物质的简单识别类似。 英文
当抗原与T淋巴细胞或B淋巴细胞表面相应的受体结合时,淋巴细胞至少会发生下列三个事件中的一种:第一种是在某些免疫应答中,淋巴细胞在抗原刺激下会分裂并分化为效应细胞(即被诱导产生免疫应答);第二种是这些淋巴细胞可能对这种抗原产生耐受或麻痹,以至于下次遇见该抗原时不能发生免疫应答,目前尚不清楚这些淋巴细胞是被杀死了还是仅仅在某种程度上失活了;第三种是淋巴细胞对于抗原的刺激不产生任何反应。此外,如果动物对某种抗原做出过免疫应答,那么当其再次接触此抗原时,通常会产生更快速、更强烈甚至有时会是性质改变了的免疫应答。这种针对同一特定抗原的免疫反应改变了的状态被称为免疫记忆。这种记忆可能既包括克隆扩增(即未经免疫的淋巴细胞通过分裂,使得再次接触同一抗原时,能产生应答的效应细胞数增加),也包括未经免疫的淋巴细胞分化为记忆细胞 [1] 。但目前尚不清楚记忆细胞只是“退休”的效应细胞——比效应细胞处于更早的分化阶段,还是通过一个独立的记忆通路分化出来的细胞。 英文
当淋巴细胞与抗原接触之后,这个淋巴细胞的“决定”是“激活”、“耐受”还是忽略主要取决于抗原的性质和浓度,还取决于它们与其他淋巴细胞和巨噬细胞的复杂的相互作用。尽管大多数的免疫原都可以激发T细胞和B细胞产生免疫应答,但是有些抗原,尤其是那些具有相同重复的抗原决定簇并且不容易在体内被代谢的抗原——所谓的“胸腺非依赖性抗原”(如肺炎球菌的荚膜多糖、大肠杆菌内毒素、聚乙烯吡咯烷酮)——首要激活B细胞(有关综述见参考文献1),其他抗原则优先激活T细胞 [48] 。通常来讲,相比B细胞而言,T细胞可以对更低浓度的抗原做出应答,而且尽管过高或过低浓度的抗原(分别为高、低耐受区间)都可以引发T细胞的耐受性,但是B细胞只会对高浓度的抗原产生耐受性 [49] 。虽然人们并不清楚抗原和受体的相互作用是如何向淋巴细胞传递信号的,但是这个过程可能包含了膜上结合的受体的构象变化和(或)其在细胞表面的重新分布(例如聚集成斑或定位在一极聚集成帽状 [50] )。 英文
T细胞和B细胞最大的区别在于它们在免疫应答中的功能不同。当B细胞被抗原激活后,它会分裂并分化为含有丰富内质网的母细胞,且部分母细胞会继续分化为浆细胞。这些细胞大多存在于淋巴组织中,并分泌大量的抗体进入血液循环。有很多方法可以用来检测分泌抗体的细胞,其中最常见的是溶血空斑实验。该实验的原理是在补体存在的情况下,单个B细胞分泌的抗红细胞抗体可以使红细胞发生溶血,从而在每个B细胞周围形成一个空斑。抗体连同各种辅助细胞(如巨噬细胞、肥大细胞、嗜碱性粒细胞)以及特殊的血清酶(如补体成分)参与了一系列超敏反应以及抵抗多种病原体的保护性免疫。此外,抗体还可以调节T细胞和B细胞的功能,通过与淋巴细胞表面的受体竞争结合抗原决定簇、将抗原转移出淋巴组织或者通过形成耐受性抗原-抗体复合物 [51] ,抗体可以抑制淋巴细胞的免疫应答;而通过将抗原定位于适当的淋巴组织或通过形成免疫性的抗体-抗原复合物,抗体可以增强淋巴细胞的免疫应答。B细胞自身也可能(但不确定)直接参与抗原转运(可能通过黏附在Fc或B细胞补体受体上的抗原-抗体±补体复合物来实现)和(或)杀死被抗体覆盖的靶细胞 [52] 。 英文
当T细胞被抗原激活后,它们会增殖、分化为母细胞,但不会产生发达的内质网也不会成为可以分泌抗体的细胞。然而,它们会分泌诸如迁移抑制因子(MIF)、趋化因子、细胞毒素、促有丝分裂因子等各种非抗原特异性因子(“淋巴因子”)。至少部分因子可能在T细胞起主要作用的细胞免疫应答中发挥作用 [53] 。但是人们尚不完全了解这些因子的化学本质、它们之间的相互关系、它们的重要性和作用机制。细胞免疫应答包括迟发性超敏反应、接触过敏、对外来组织的排斥、移植物抗宿主反应(移植的外来T淋巴细胞对受体抗原的反应,经常会导致受体死亡)以及机体对各种微生物的免疫力。在所有这些免疫应答中,T细胞要发挥功能都需要巨噬细胞(可能是通过分泌淋巴因子的方式)的参与,并且巨噬细胞通常在反应位点扮演主要的角色 [54] 。在体外实验中,T细胞也可以通过分裂、分泌淋巴因子、杀伤靶细胞或者维持病毒复制(有关综述见参考文献1)等方式对抗原产生应答。尽管越来越多的证据表明,在某些环境下T细胞可以成为“杀手细胞” [55] ,但是关于T细胞自身是否可以直接杀伤靶细胞还是必须通过激活其他细胞(如巨噬细胞)杀伤靶细胞目前尚有争论。 英文
尽管T细胞本身一般不分泌抗体,但现在已经知道它们在帮助B细胞对免疫原作出应答方面发挥着重要的作用。因此,在这类免疫应答中,T细胞被称为“辅助”细胞,而B细胞则被称为“抗体–形成前体”细胞。关于T-B细胞协作的首例直接证据是在1966年发现的。研究人员发现,向经放射照射的小鼠同时移植胸腺细胞和骨髓细胞,其产生抗绵羊红细胞(SRBC)抗体的能力比那些只移植胸腺细胞或骨髓细胞的小鼠产生抗体的能力强的多 [56] 。后来人们发现,在这类实验中,所有抗体分泌细胞(也就是产生抗SRBC抗体的细胞)都来自于移植的骨髓细胞 [57] 。用化学成分确定的抗原进行的独立研究显示,在抗体免疫应答中,T-B细胞的协作包括T细胞对免疫原上的某一抗原决定簇进行识别,并且帮助B细胞对同一免疫原上不同抗原决定簇产生反应 [58] 。尽管目前已经清楚这种协作通常是通过T细胞和B细胞受体之间的“抗原桥”来介导的,但人们尚不清楚这个桥是介于T细胞和B细胞之间还是介于脱落的T细胞受体(可能被巨噬细胞摄取后呈递在其表面)与B细胞之间。也不清楚这个桥的作用是以特定的免疫原形式(如浓缩或多价)向B细胞“呈递”抗原,还是把B细胞与T细胞或第三方细胞(如巨噬细胞)拉到一起,以便于非特异性的、小范围内起作用的因素(如化学媒介或细胞膜-细胞膜相互作用)在它们之间发挥作用(图2)。尽管已经证明T细胞可以分泌一些能够加强B细胞免疫应答的非特异性因子 [59] ,但是人们并不清楚这些因子在正常的T-B细胞协作中发挥着什么样的作用。最近有证据表明,至少在体外免疫应答中,T-B细胞协作可能与T细胞分泌一种具有抗原特异性的IgM样因子(受体?)有关。这种因子可以与抗原结合形成复合物,然后会被巨噬细胞吞噬 [60] 。 英文
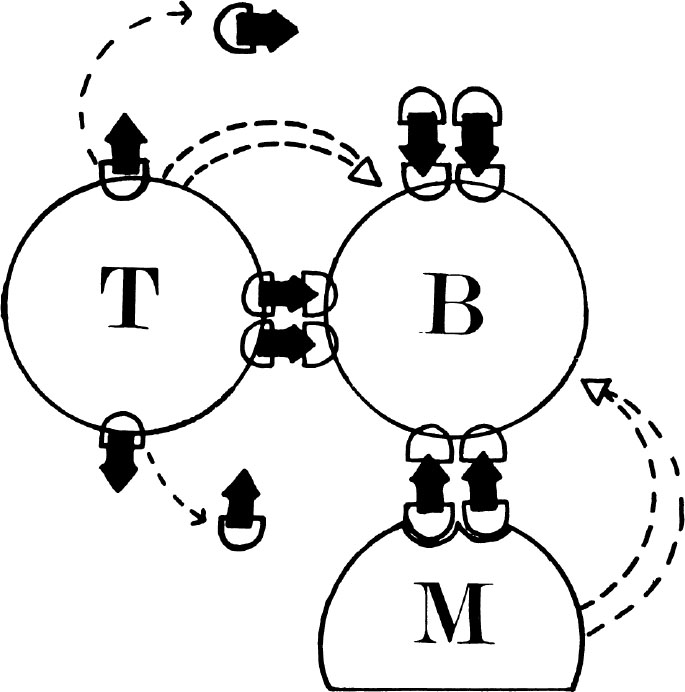
图2. 体液免疫应答中T-B细胞协作的可能机理。T细胞和B细胞受体之间的“抗原桥”(→),可能发挥如下作用:(i)在T细胞表面将抗原呈递给B细胞;或者作为释放T细胞受体-抗原复合物的基体,这种复合物可以是游离的,也可以是位于第三方细胞如巨噬细胞表面的。(ii)把B细胞和T细胞或第三方细胞拉到一起,从而使小范围内起作用的因素在它们之间发挥功能。
在没有T细胞辅助的情况下,有一些抗原(“胸腺非依赖性抗原”)可以刺激部分B细胞分泌IgM抗体(有关综述见参考文献1)。这表明T-B细胞的协作并不总是抗体产生所必需的。不过发现体液免疫中T细胞与B细胞的协作仍然是免疫学研究中的一个重大进展,它解释了之前提出的缺乏T细胞的动物为什么会出现抗体反应受损这一问题。最近还有间接证据表明T细胞不但可以抑制B细胞的应答,也可以增强B细胞的活性 [61] 。同时,它们还可以增强 [62] 或抑制 [63] 其他T细胞的功能。人们并不清楚这些相互作用是否包括相互作用的细胞之间受体的抗原桥连接。总之,考虑所有这些分泌的抗体对T细胞和B细胞功能的增强或抑制作用,我们可以想象免疫系统是高度复杂且精密调控的,在这个系统中每种细胞和免疫应答都可以调控另一种。 英文
随着我们的认识,免疫系统中存在着两种不同来源、不同性质、不同免疫功能的淋巴细胞,它们相互调控着彼此的活性,免疫学研究已进入一个新时代。在健康和疾病方面,对免疫系统功能的深入了解使我们有可能通过合理的实验,选择性地控制不同免疫细胞类型及其多样应答,以改善被传染病、自身免疫疾病、癌症、免疫缺陷疾病或器官移植所困扰的患者的生活。另外,现代免疫学也提供了许多可用于研究多种生物学问题(包括分化、遗传控制、细胞间相互作用以及细胞膜受体–配体间相互作用)的有效模型和强大工具。 英文
非常感谢格里夫斯、米奇森和欧文宝贵的建议。此外,参考文献引用可能不全面,仅作为查阅相关文献的一个途径。 英文
(张锦彬 翻译;秦志海 审稿)