
下载掌阅APP,畅读海量书库
立即打开



 T and B Lymphocytes and Immune Responses
T and B Lymphocytes and Immune Responses
M. C. Raff
Editor’s Note
In this paper, Canadian-born cell biologist Martin C. Raff sums up what was then known about T and B lymphocytes. The two white blood cell types are found in peripheral lymphoid organs where they look identical, and Raff had previously identified a T cell marker, an achievement that won him immediate international recognition. T and B cells have different origins, properties and immunological functions which modulate each other’s activities. Their discovery, Raff says, marks a new era in immunology in which powerful research tools and accessible models are likely to light on biological issues as well as disease. 中文

The recognition of two distinct classes of lymphocytes has been a turning point in immunology. Immunological models and tools may help to provide the answers to many biological problems. 中文

IMMUNOLOGY has become an exciting science of its own. Nonetheless, what is being learned about lymphocytes and the immune responses that they mediate has important implications for medicine and other branches of biology. Unfortunately, the private language of immunology has made it difficult for non-immunologists to join in the excitement. This article attempts to review what is known in general terms about the cellular basis of immunity. (For a more detailed review of lymphocytes and their roles in immune responses, see ref. 1. 中文
Immunology is concerned with the specific responses an animal makes when foreign materials (antigens or immunogens) are introduced into its body. Such immune responses are made by all vertebrates and consist of the production of specific immunoglobulin protein molecules (antibodies) and/or specifically reactive cells, both of which can circulate in the blood and react specifically with antigen. As a result of this reaction, the foreign material may be inactivated (for example, bacterial toxins), killed (for example, infecting organisms or transplanted cells) and/or phagocytosed by cells of the reticuloendothelial system. On the other hand, in some cases, such immune responses may have deleterious effects on the host, such as in hypersensitivity reactions (hayfever and drug allergy, for example), where antigen reacting with antibody fixed to basophils and mast cells causes the release of histamine and other pharmacological mediators of inflammation. In general, immune responses which can be transferred to another animal by means of serum from a sensitized donor (containing antibody) are termed humoral immune (or antibody) responses, whereas those that can be transferred by sensitized cells but not by serum are called cell-mediated immune responses. 中文
While immunochemists were unravelling the structure of antibody in the 1950s and early 1960s, cellular immunologists were demonstrating that lymphocytes are the principal cells involved in immune reactions. The most convincing experiments were those showing that relatively pure populations of rat lymphocytes obtained from the chief lymphatic vessel, the thoracic duct, could transfer both cellular and humoral immunity to irradiated rats, which could not respond immunologically themselves as their lymphocytes had been killed by the radiation (reviewed in ref. 2). In addition, depleting animals of lymphocytes by prolonged drainage of the thoracic duct was found to impair their immune responsiveness 2 . Thus lymphocytes, whose origins and functions had been a mystery for so long, were established as “immunocompetent” cells. 中文
It was soon realized that lymphocytes are not a homogeneous population. Several lines of evidence suggested that there are two distinct types of immunocompetent lymphocytes: one which requires the thymus gland for development and is responsible for cell-mediated immunity and another which develops independently of the thymus and mediates humoral antibody responses. The evidence came from studies in birds, rodents and man in the 1960s. In birds 3,4 and rodents 5 it was found that removing the thymus from an embryo or newborn markedly impaired the cell-mediated immune responses of the animals when they grew up, but had much less effect on humoral immunity. On the other hand, removal at hatching of the bursa of Fabricius 3,4 , a cloacal lymphoid organ unique to birds, impaired the bird’s ability to make antibody, but had little effect on cell-mediated immunity. Investigations of patients with immunological deficiency diseases also showed that humoral and cell-mediated immunity could be separately affected (reviewed in ref. 6): patients with Bruton-type congenital agammaglobulinaemia could not make antibody and were deficient in lymphoid cells producing antibody, but had normal cell-mediated immunity, whereas children with congenitally hypoplastic thymus glands (for example, Di George’s syndrome) had markedly impaired cell-mediated immunity but could make relatively normal amounts of antibody in response to some antigens. 中文
In the past few years the two-lymphocyte model of immunity has been firmly established (at least in birds and mammals), with two “central” lymphoid organs—the bursa, or its mammalian equivalent (still unidentified), and the thymus—producing lymphocytes independently of antigen, and seeding them out to the “peripheral” lymphoid organs (that is, lymph nodes, spleen and gut-associated lymphoid tissues) where they await contact with antigen which will induce them to differentiate into “effector” cells (see later). In the peripheral lymphoid tissues the lymphocytes derived from thymus are referred to as T cells, while those derived from the bursa in birds, or its equivalent in mammals, are called B cells 7 . 中文
Until recently it was thought that specific immune responses were confined to vertebrates. There is now evidence, however, that some invertebrates, such as annelids and tunicates, can reject foreign tissues and that these primitive immunological responses can display specificity and possibly short-term memory 8 (that is, an increased and/or faster response on second exposure to the same antigens). These reactions are mediated by macrophage-like cells (coelomocytes) and possibly by soluble effector molecules having relatively little specificity 8 . As there is no evidence that invertebrates have lymphocytes or immunoglobulins, it seems likely that specific cellular immunity evolved before the appearance of these two principal mediators of vertebrate immunity. 中文
All vertebrates have lymphocytes and probably thymus tissue (at least at some stage in their development) and are capable of producing antibody and cell-mediated immune responses 8 . Lower vertebrates (lampreys and hagfish, for example) have little organized lymphoid tissue and can produce only one class (IgM-like) of antibody. Rudimentary lymph node-like structures are first found in Amphibia which make two classes of antibody. Birds are the first vertebrates in which a clear dichotomy of the lymphoid system has been established, and are unique in having two discrete central lymphoid organs, thymus and bursa, producing T and B lymphocytes respectively. Mammals have abundant and highly organized lymphoid tissues, can elaborate a variety of different classes of antibody (such as IgG, IgM, IgA, IgE, IgD in man) and have distinct T and B lymphocyte populations, although the site of B cell development is still uncertain. It is not known whether vertebrates below birds have separate classes of T and B cells. 中文
In most animals, lymphocytes first appear in the foetal thymus. The thymus anlage is composed of epithelial cells and is derived from the third and fourth pharyngeal pouches. Although in the past it had been suggested that thymus lymphocytes (thymocytes) develop from thymus epithelial cells, experiments in chickens and mice have clearly established that haemopoietic stem cells from foetal yolk sac and liver migrate into the thymus anlage and there proliferate and differentiate into thymus lymphocytes, presumably under the inductive influence of the thymus epithelium 9 . In mice (gestation 20 days) the first stem cells, which seem to be large basophilic blast-like cells, arrive in the thymus around day 11, and the first small lymphocytes are seen by day 15 or 16 of embryonic life 9 . Using radioactive 10 , chromosome 5,11 and surface antigenic 9 markers, it has been shown that lymphocytes migrate from thymus to peripheral lymphoid tissues to make up the T lymphocyte population. Although this begins just before birth in mice, most of the seeding occurs in the first week of life 9 . Therefore, if the thymus is removed in the first days of life the mouse will grow up with a marked deficiency of T cells and thus impaired cell-mediated immunity, whereas thymectomy done later in life has much less effect 5 . In adult animals, stem cells from bone marrow migrate to thymus, and thymus lymphocytes continue to seed to the periphery, but these processes take place at a much reduced rate by comparison with the foetus and newborn 5,11 . 中文
Most thymus lymphocytes are immunologically incompetent (that is, they cannot respond to antigen) and differ in other ways from peripheral T cells, suggesting that there is another differentiation step from thymocyte to T lymphocyte. Recently it has been demonstrated that there is a small subpopulation (~2 to 5%) of thymus cells, located in the thymus medulla, which is immunologically competent and has most of the properties of peripheral T lymphocytes 9,12,13 . This suggests that the second differentiation step may occur within the thymus and that T cell development may be visualized as stem cell→thymocyte→“mature” thymus lymphocyte→peripheral T lymphocyte (Fig. 1). This scheme is almost certainly an oversimplification, however, for there is some evidence that cells may leave the thymus at varying stages of maturation, or perhaps as distinct cell lines, giving rise to subpopulations of peripheral T cells with different properties and functions 13 . In addition, the role of putative thymus humoral factors or hormones (thymosin, for example) is still unclear, although there is evidence that they probably do not induce stem cells to differentiate to lymphocytes outside the thymus, but may influence peripheral T cells in some way 14 . 中文
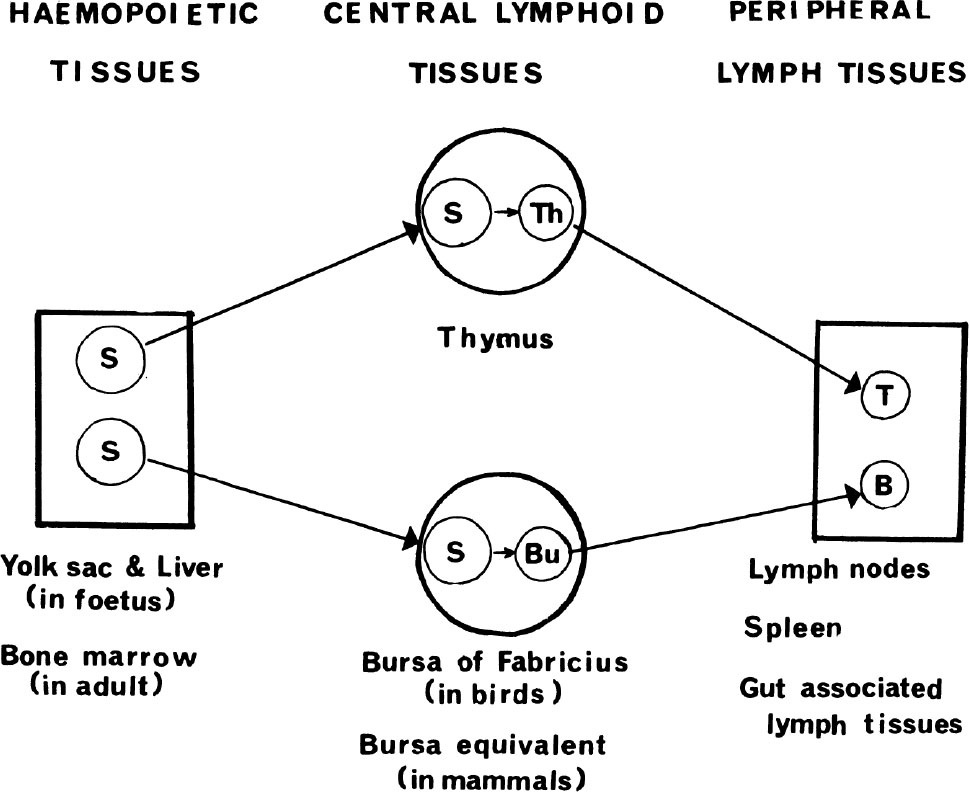
Fig. 1. Diagrammatic (and oversimplified—see text) representation of T and B lymphocyte development showing migration of stem cells (S) to thymus and bursa where they differentiate to thymus (Th) and bursal (Bu) lymphocytes, some of which migrate to the peripheral lymphoid tissues as T and B lymphocytes respectively.
In birds, B cell development is dependent on the bursa of Fabricius which arises as a sac-like evagination of the dorsal wall of the cloaca on day 5. Chromosome marker studies have shown that stem cells (morphologically identical to those seen in the foetal thymus) begin to migrate from yolk sac to the bursa around days 12 to 13 and there differentiate to lymphocytes within 1 or 2 days 9 . By day 14, bursa lymphocytes with IgM on their surface can be seen, and bursa lymphocytes bearing IgG are seen a few days later 15 . The migration of bursal lymphocytes to peripheral lymphoid tissues has been demonstrated by isotope labelling experiments. Embryonic bursectomy results in marked depletion of peripheral B lymphocytes and a marked impairment in antibody (that is, immunoglobulin) production 15 . Recently it has been found that injecting anti-μ antibody (that is, specific for the heavy chains of IgM) before hatching, combined with neonatal bursectomy, suppresses later production of IgG as well as IgM 15 . This suggests that even B cells that will eventually produce IgG initially express IgM on their surface, and is strong evidence for an IgM→IgG switch within individual B cells. Whether this switch is driven by antigen, as suggested by experiments in mice 16 , or occurs independently of antigen stimulation, as suggested by experiments in chickens 15 , is unsettled. 中文
In mammals, it is still not clear where stem cells differentiate to B-type lymphocytes, although it is known not to be in the thymus. It has been suggested that gut-associated lymphoid tissues (like Peyer’s patches, tonsils, appendix, and so on) may serve as “bursa-equivalent”, but there is little evidence to support this. In rodents, at least, there is increasing evidence that lymphocytes are produced in large numbers in the haemopoietic tissues themselves 17 (that is, liver in embryos and bone marrow in adults) and it seems likely that these tissues not only supply the stem cells for both T and B cell populations but are also the sites where stem cells differentiate to B-type lymphocytes. 中文
It is not clear at what stage stem cells are committed to becoming lymphocytes or to becoming T or B cells. The finding of multipotential haemopoietic stem cells (that is, cells capable of becoming any of the mature blood cell types, lymphoid or myeloid) in early mouse embryonic thymus 18 suggests that commitment may not occur until stem cells enter the microenvironment of the thymus or bursa (or bursa equivalent). 中文
As resting T and B lymphocytes are morphologically indistinguishable and are found together in all peripheral lymphoid tissue, it has been essential to find ways of distinguishing and separating them in order to study their individual properties. The demonstration of important surface differences between them has been particularly useful in this regard. Some of these surface differences can be recognized by antibody 19 . For example, the θ alloantigen (defined by alloantibody made in one strain of mouse against thymocytes of another strain) is present on mouse thymocytes and T cells, but absent from B lymphocytes, and this has proved to be a convenient surface marker for T cells in mice 19 . On the other hand, readily demonstrable surface immunoglobulin (Ig) (refs. 20, 21) and the heteroantigen, “mouse-specific B lymphocyte antigen”(MBLA) (ref. 19)—defined by hetero-antibody made in rabbits against mouse B cells—can serve as B cell markers. With antisera reacting specifically with the surface of one or other lymphocyte type, either cell population can be killed in the presence of complement, and thus eliminated from a cell suspension. Alternatively, one can use antibody on digestible solid-phase immunoabsorbents 22 , or fluoresceinated antibody and fluorescence-activated electronic cell sorting 23 , to purify either type of cell. In addition to surface antigenic differences between T and B cells, the latter can bind antibody-antigen-complement complexes by means of surface complement receptors 24 , and antibody-antigen complexes by means of receptors for the Fc part of complexed Ig 25 ; resting T cells do not have these receptors. The functions of Fc and complement receptors on B cells are unknown, but it has been suggested that they may be important in antigen localization in the lymphoid tissues, in B cell activation by antigen and/or in putative killing by B cells of target cells coated with antibody. 中文
Most T lymphocytes continuously recirculate between blood and lymph, passing out of the blood through specialized post-capillary venules in lymph nodes and Peyer’s patches, passing through the substance of the lymphoid tissues and entering the efferent lymph; they then re-enter the bloodstream by way of the thoracic duct 2,5 . Although most B lymphocytes seem not to recirculate, some apparently do, but through different areas of the lymphoid tissues and with a slower transit time than T cells 26 . In the peripheral lymphoid tissues, T and B cells are found in more or less separate areas, the so-called thymus-dependent areas (periarteriolar sheath of spleen, paracortex of lymph nodes, and interfollicular areas of gastrointestinal lymphoid tissues) and thymus-independent areas (lymph follicles and peripheral regions of splenic white pulp, follicles and medulla of lymph nodes and follicles of gastrointestinal lymphoid tissues) respectively 27 . When radiolabelled T or B cells are injected into an animal, they migrate specifically to their respective areas 27 . Although both T and B lymphocyte populations are heterogeneous 1 , T cells have a longer generation time 28 on average and are slightly larger 29 , more dense 24 , less adherent 24 (to various materials such as glass, plastic, nylon, and so on) and more negatively charged than B cells 30 . In addition, T lymphocytes are preferentially depleted by anti-lymphocyte serum 31 (which acts principally on recirculating cells), but in general are less sensitive to cytotoxic drugs (for example, cyclophosphamide 32 ), corticosteroids 33 and irradiation 34 . T and B cells also differ in their in vitro responses to a variety of “mitogens”, such as plant extracts (phytomitogens), bacterial products (like endotoxin) or antibodies to lymphocyte surface antigens, which stimulate a relatively large proportion of T and/or B lymphocytes to divide and differentiate into blast cells. Although pokeweed stimulates both T and B cell proliferation, concanavallin A (Con A), phytohaemag-glutinin (PHA) and lentil stimulate only T cells, and lipopolysaccharides (for example, E. coli endotoxin) and anti-Ig sera stimulate only B cells 35 . It is of interest that although soluble Con A and PHA selectively activate T cells, they bind equally well to B cells, and if covalently linked to solid-phase materials they stimulate B cell proliferation 35 . Mitogen stimulation of lymphocytes is being intensively studied as a possible model of lymphocyte activation by specific antigen. These studies have made it clear that there is more to lymphocyte activation than simple binding of ligand to surface receptors. 中文
The central dogma of immunology is the clonal selection hypothesis which suggests that at some time in ontogeny and independently of antigen, individual lymphocytes (or clones of lymphocytes) become committed to responding to one, or a relatively small number of antigens; they express this commitment through antigen-specific receptors on their surface. Thus, when an antigen is introduced into the body it selects out those lymphocytes which already have receptors for the antigen on their surface; the interaction of antigen with receptors initiates the activation of the specific cells. There is now an impressive body of evidence supporting the clonal selection hypothesis for both T and B lymphocytes. Thus T and B cells have been shown to bind antigen to their surface 36 (although it has been more difficult to demonstrate T cells binding antigen than B cells) and in general only a small proportion of lymphocytes (~1 in 10 4 to 10 5 in unimmunized animals) bind any one antigen. Furthermore, if lymphocytes are exposed to a highly radioactive antigen, both T and B cell responses to that antigen can be selectively abolished, while responses to other antigens are unaffected 37 . Similarly, B cells capable of responding to a particular antigen specifically adhere to glass beads coated with the antigen and can thus be specifically removed from a cell suspension 38 . Although T cells tend not to adhere under these conditions 38 for reasons that are unclear, T cells responsive to cell surface alloantigens can be selectively removed in cell monolayers bearing the specific alloantigens 39 . 中文
In 1900, Ehrlich proposed that cells producing antitoxins (now known to be B cells) had antitoxin molecules as receptors on their surface. The more recent version of the receptor hypothesis suggests that B lymphocytes have antibody molecules (that is, Ig) as receptors for antigen, which, at least in their combining sites, are identical to the antibody which the cell or its progeny will eventually secrete. There is now good evidence for this view, in that B cells have been shown to have Ig molecules on their surface (~10 4 to 10 5 a cell) (refs. 20, 40) and anti-Ig antibody inhibits their ability to bind or respond to antigens (reviewed in ref.1). There is also increasing evidence that the antigen-specificity of receptors and secreted antibody are the same for any one B lymphocyte clone 41,42 . The Ig class of the receptors and that of the ultimately secreted antibody may not, however, always be the same, for B cell precursors of some IgG secretory cells seem to have IgM receptors 15,16 . As different antibody classes (for example, IgG and IgM) seem to be able to share the same specificity (that is different Ig constant regions can be associated with identical Ig variable regions 43 ) and IgM→IgG switch within a single clone need not imply a switch in specificity. In mice, at least, there is some evidence that most virgin B cells have IgM receptors (in its 7-8S monomeric form 44 ) which may switch class after a primary exposure to antigen 16 . The more fundamental question of how antibody diversity is generated, that is how an animal develops the ability to synthesize such a large number of different Ig molecules (receptors and secreted antibodies) is still being debated. Germ-line theories, which suggest that one is born with a large number of variable region Ig genes, are competing with various somatic theories, which postulate that one is born with few variable region Ig genes and that some somatic process (for example, mutation or recombination) creates a large number. 中文
The chemical nature of receptors on T cells is probably the most controversial issue in cellular immunology at present. The simplest and most logical view, that only antibody can recognize antigen and that all antigen-specific receptors must be Ig, has been challenged by the failure of many investigators to demonstrate Ig directly on the surface of T cells, or to inhibit various T cell responses with anti-Ig sera. Indeed, there is now growing support for the idea that surface components other than classical immunoglobulin may play an important role in T cell recognition of and/or response to at least some antigens. The principal candidates for such T cell “receptors” are the products of the immune response (Ir) genes that are genetically linked to the chief histocompatibility loci 45 . These Ir genes influence T cell responses to a variety of antigens 46 . The exquisite specificity of T cell responses, which resembles very closely the specificity of antibody and B cell recognition 47 , taken together with the various (but still controversial) demonstrations of Ig on T cells (reviewed in ref. 1), makes one reluctant, however, to give up the idea that T cells have Ig receptors. It is possible that T cells (and possibly B cells) have at least two “recognition” systems, one involving Ig and another mediated by Ir gene products, the general importance of each varying depending on the antigen, the response and/or the subclass of T cell. The putative non-Ig recognition system could be analogous to the primitive recognition of foreignness seen in invertebrates. 中文
When an antigen combines with its corresponding receptors on a T or B lymphocyte, one of at least three things can happen to the lymphocyte: first, it may be stimulated to divide and differentiate to become an effector cell in some type of immune response (that is, it is induced to respond immunologically); second, it may become immunologically tolerant or paralysed, so that it will not be able to respond the next time antigen is given; it is not known if such cells are killed or simply inactivated in some way; third, it may be unaffected by the encounter. In addition, if the animal makes an immune response to the antigen, on subsequent exposure to the same antigen, it will usually give a faster, greater and sometimes qualitatively different response. This altered state of immune reactivity to a specific antigen is called immunological memory. It is likely that memory involves both clonal expansion (that is, division of virgin lymphocytes to give an increased number of cells able to respond on second exposure) and differentiation of virgin cells to memory cells 1 , but it is unclear whether memory cells are simply retired effector cells, cells at an earlier stage of differentiation than effector cells, or are derived by differentiation along a separate memory pathway. 中文
The “decision” of an individual lymphocyte on encounter with antigen—whether to “turn-on”, “turn-off” or ignore—depends largely on the nature and concentration of the antigen, and upon complex interactions with other lymphocytes and with macrophages. Although most immunogens can stimulate both T and B cell responses, some, particularly those with repeating identical determinants and which are poorly catabolized—the so-called “thymus-independent antigens”(for example, pneumococcal polysaccharide, E . coli endotoxin, polyvinylpyrrolidone)—chiefly stimulate B cells (reviewed in ref.1), whereas others preferentially activate T cells 48 . In general, T cells respond to lower concentrations of antigen than do B cells, and although T cells may be paralysed at very low and very high concentrations of antigen (low and high zones of tolerance respectively) B cells seem to be paralysed only at high antigen concentrations 49 . The way in which the antigen-receptor interaction signals a lymphocyte is unknown, although it probably involves allosteric changes and/or redistribution (for example, aggregation into patches or localization over one pole–cap formation 50 ) of the membrane-bound receptors. 中文
The most important differences between T and B cells concern their different functions in immune responses. When B cells are activated by antigen they divide and differentiate into blast cells with abundant endoplasmic reticulum, and some go on to become plasma cells. These cells remain in the lymphoid tissues for the most part and secrete large amounts of antibody which circulates in the blood. Individual antibody-secreting cells can be detected by a variety of techniques, the most common being the plaque-forming cell assay, in which anti-erythrocyte antibody released from single B cells lyses erythrocytes in their immediate environment in the presence of complement. Antibodies, in conjunction with various accessory cells (macrophages, mast cells and basophils, for example) and particular serum enzymes (complement components, for example), are responsible for a variety of hypersensitivity reactions and protective immunity against many pathogenic organisms. In addition, antibody serves to regulate the function of both T and B cells, inhibiting their responses by competing with lymphocyte receptors for the antigenic determinants, diverting antigen from the lymphoid tissues or by forming tolerogenic antibody-antigen complexes 51 , and enhancing responses by localizing antigen to appropriate lymphoid tissues or perhaps forming immunogenic antibody-antigen complexes. It is also possible (but not established) that B cells themselves play a direct part in transporting antigen (perhaps as antigen-antibody ± complement complexes adhering to Fc or complement receptors on B cells) and/or in killing target cells with coated antibody 52 . 中文
When T cells are activated by antigen, they proliferate and differentiate to become blast cells, but they do not develop significant amounts of endoplasmic reticulum and do not become antibody-secreting cells. They do, however, secrete a variety of non-antigen-specific factors (“lymphokines”) such as migration inhibition factors (MIF), chemotactic factors, cytotoxic factors and mitogenic factors, at least some of which presumably play a role in cell-mediated immune responses, for which T cells are primarily responsible 53 . The precise chemical nature of these factors, the relationship between them, their significance and mechanisms of action are, however, incompletely understood. Cell-mediated immune responses include delayed hypersensitivity, contact sensitivity, rejection of foreign tissues, graft versus host responses (where injected foreign T lymphocytes respond against the antigens of the recipient, often resulting in recipient death) and immunity to various microbes. In all of these responses, T cells enlist the help of macrophages (probably through the secretion of lymphokines). The latter are usually the predominant cells at the site of these reactions 54 . T cells can also be demonstrated to respond to antigen in vitro by dividing, secreting lymphokines, killing target cells, or supporting viral replication (reviewed in ref.1). Whether T cells themselves can directly kill target cells, or do so only by activating other cells (such as macrophages) is still controversial, although there is increasing evidence that they can become “killer cells” under some circumstances 55 . 中文
Although T cells do not themselves secrete antibody in the usual sense, it is now known that they play an important role in helping B cells to make antibody responses to most immunogens. Thus, in these responses T cells are referred to as “helper” cells, and B cells as “antibody-forming precursor” cells. The first direct evidence for such T-B cell cooperation was provided in 1966 by the observation that irradiated mice given both thymus cells and bone marrow cells made a far greater antibody response to sheep erythrocytes (SRBC) than recipients of either thymocytes or bone marrow cells alone 56 . Subsequently it was shown that all of the antibody-secreting cells (that is, those making anti-SRBC antibody) in this type of experiment came from the bone marrow inoculum 57 . Independent studies with chemically defined antigens showed that T-B cell cooperation in antibody responses involved T cells responding to one antigenic determinant on an immunogen and helping B cells to respond to different determinants on the same immunogen 58 . Although it is clear that cooperation is usually mediated by such an “antigen bridge” between T cell and B cell receptors, it is still uncertain whether the bridge is between T and B cells themselves, or between shed T cell receptors (perhaps taken up on the surface of macrophages) and B cells, and whether the bridge serves to “present” antigen to B cells in a particularly immunogenic form (concentrated and multivalent, for example) or to bring B cells close to T cells or a third party cell (such as macrophage) so that a nonspecific, short-range factor (for example, chemical mediator or membrane-membrane interaction) can operate between them (Fig. 2). Although it has been shown that T cells can secrete non-specific factors which can enhance B cell responses 59 , their role in normal T-B cell cooperation is still uncertain. There is recent evidence that, in some in vitro responses at least, cooperation may involve the release by T cells of antigen-specific IgM-like factors (?receptors) complexed with antigen, which are subsequently taken up on macrophages 60 . 中文
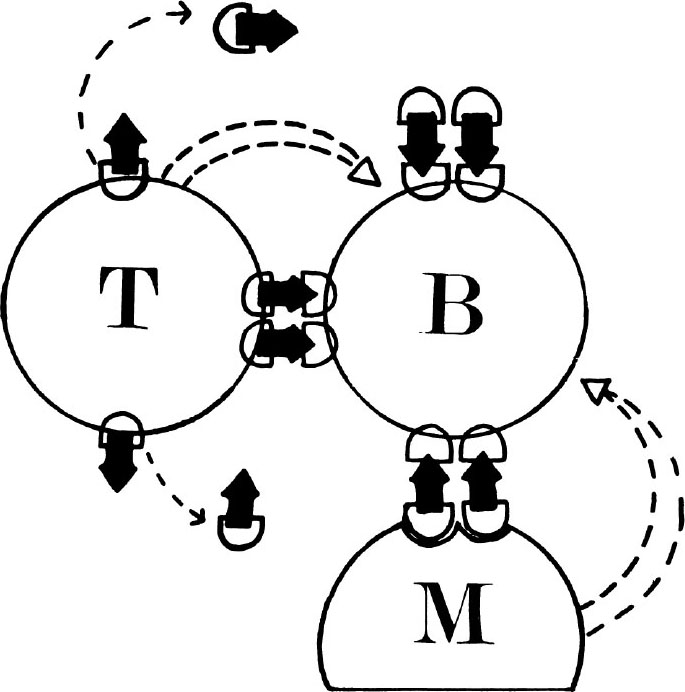
Fig. 2. Possible mechanisms of T-B cell collaboration in humoral antibody responses. The “antigen bridge”(→) between T and B cell receptors could serve to : (i) present antigen to B cells on the surface of T cells or as a matrix of released T cell receptors complexed with antigen, either free or on the surface of third party cells such as macrophages, or (ii)bring B cells together with T cells or a third party cell so that a short-range factor can operate between them.
There are antigens (“thymus-independent antigens”) which seem to be able to stimulate at least some B cell clones to secrete IgM antibody without the help of T cells (reviewed in ref. 1), suggesting that T-B cell collaboration is not always essential for antibody production. Nonetheless, the discovery that T cells cooperate with B cells in humoral immunity has been an important advance and has explained the previous paradox of impaired antibody responses in T cell deficient animals. There is recent indirect evidence that T cells can inhibit B cell activity as well as enhance it 61 , and that they can enhance 62 and inhibit 63 the functioning of other T cells. It is not known if these interactions involve antigen bridging between the receptors of the interacting cells. Taken together with the enhancing and inhibiting effects of secreted antibody on both T and B cell functions, a picture is emerging of a highly complex and finely controlled immune system, with each type of cell and response modulating the others. 中文
With the recognition that there are two distinct classes of lymphocytes with different origins, properties and immunological functions which modulate each other’s activities, the door has opened to a new era of immunology. The resulting insight into the functioning of the immune system in health and disease has paved the way for rational attempts to manipulate selectively the different cell types and their various responses for the benefit of patients with infection, autoimmune disease, cancer, immune deficiency states, and organ grafts. And present day immunology provides a number of readily accessible models and powerful tools for studying a variety of biological problems, including differentiation, genetic control, cell interactions, and membrane receptor-ligand interactions. 中文
I am grateful to M. F. Greaves, N. A. Mitchison and J. J. T. Owen for helpful discussion. The bibliography, which is incomplete, is meant only as an arbitrary way into the relevant literature. 中文
( 242 , 19-23; 1973)
Martin C. Raff
Medical Research Council Neuroimmunology Project, Zoology Department, University College, Gower Street, London WC1
References: