
下载掌阅APP,畅读海量书库
立即打开



 哺乳动物细胞核内的DNA复制位点
哺乳动物细胞核内的DNA复制位点
休伯曼等
编者按
在DNA结构发表近20年后,分子生物学家们仍然在孜孜不倦地研究作为遗传物质的DNA在高等生物活细胞(真核细胞)中是如何复制的。这篇文章的意义在于,它显示了科学家们对于DNA的复制过程的认识仍然浅显。休伯曼等人研究了海拉(HeLa)细胞(一种源自人体癌细胞的细胞株)的有丝分裂过程。“S”期是指即将分裂的细胞积累分裂成两个细胞所需物质的阶段。他们总结认为DNA首先在核膜附近合成,然后迁移到整个细胞核。 英文

DNA复制发生在整个细胞核内,而不是局限于核膜的内表面。 英文

雅各布、布伦纳和居赞 [1] 在关于细菌DNA复制的“复制子”模型中首次提出膜位点参与DNA的复制,但是其后这方面所积累的证据并不确凿。在原核细胞中,DNA复制点和细胞膜的共沉淀已经得到证明 [1-4] ,同时发现,此复制起点能与细胞膜结合 [5] 。从大肠杆菌中分离得到了不含膜脂的复制的DNA-蛋白复合体 [6] ,且有报道称仅大肠杆菌染色体上的复制起点与细胞膜相连,生长点与其不相连 [7] 。 英文
通过 3 H–胸腺嘧啶核苷酸(TdR)脉冲标记的哺乳动物细胞的分级分离实验发现,复制点与细胞核核膜(或者其他一些大的、轻的、疏水的细胞结构)相结合 [8-11,13] 。使用薄片电子显微放射自显影法检测脉冲标记非同步的HeLa细胞,同样发现了复制点与细胞核核膜相结合。追踪实验表明与核膜相连的标记1小时后明显转移到细胞核内部 [13] 。 英文
另一方面,大部分电子显微放射自显影实验表明,复制能在整个细胞核中发生。例如,科明斯和挂札 [14] 用 3 H-TdR脉冲标记非同步的人羊膜细胞5分钟或者更长时间,然后将其切片进行放射自显影,研究结果发现颗粒遍布整个细胞核。然而,如果把同步化为S期初期的细胞脉冲标记5或10分钟,检测到的颗粒则主要分布在核膜上。科明斯和挂札由此推断,复制的起始发生在核膜上,而复制则发生在细胞核内的任何区域。 英文
下面三个小组的研究得到的结果略有不同:布隆德尔 [15] 用脉冲处理KB细胞(一种口腔癌细胞)2分钟,威廉姆斯和奥克伊 [16] 用脉冲处理中国仓鼠细胞10分钟或者更长时间,以及厄兰森和德阿尔旺 [17] 用脉冲处理HeLa细胞15分钟。与科明斯和挂札 [14] 的结论相一致的是,上述三个研究组都认为在S期的大部分时期,颗粒存在于整个细胞核中。但是与科明斯和挂札的结论不同的是,他们发现与S期初期相比这些颗粒在S期末期出现周边模式的频率更高。凯 [18] 等人的细胞分级分离实验同样阐明了DNA与核膜的结合主要是发生在复制的后期,而不是复制的初期。 英文
不考虑更接近细胞核核膜的复制时期到底出现在S期的哪个阶段,大部分电子显微放射自显影实验得到的一个重要的结论就是,至少在S期的某些阶段内,复制发生在整个细胞核内。这一结论是如何与那些表明正在复制的DNA与核膜相结合的实验保持一致的?在短短2分钟的脉冲时间内合成大量的DNA是可能的。尽管大部分DNA的复制速率在每分钟0.5~1.2 μm,但是休伯曼和里格斯 [19] 的研究表明,中国仓鼠细胞的DNA复制速率可以达到每分钟2.5 μm。因此在2分钟的时间内可以合成5 μm的DNA。假设这些DNA是在细胞核核膜上合成,然后延伸出去,那么该DNA就可以到达直径为5~8 μm的细胞核的中心,从而造成了对DNA合成位点位置的错误判断。 英文
上述这种影响可以通过缩短脉冲时间来排除。我们计算表明,在曝光几个月以后,0.5分钟的脉冲时间能够提供足够的颗粒,其分辨能力(可分辨小于1.25 μm新合成的DNA)可以将单独发生在细胞核核膜与发生在细胞核的其他区域的复制区分开来。 英文
大部分实验中,我们使用中国仓鼠(CHO)细胞,因为这种细胞容易同步化。将这种细胞用 3 H-TdR脉冲标记0.5分钟后,进行清洗、固定、包埋、切片以及染色。切片放在铜网上,按照标准技术进行放射自显影 [20] 。曝光几个月后,显影冲洗,用电子显微镜观察铜网。 英文
图1显示了非同步培养的细胞中颗粒在细胞核中的各种分布情况。图1A和图1B的细胞展示的是整个细胞核都被标记的情况(普遍模式),而图1C和图1D的细胞展示的是被标记区域主要出现在细胞核核膜周围的情况(周边模式)。图1E中颗粒主要集中在浓缩的染色质区域,通常靠近核膜。图1F中颗粒遍布细胞的整个细胞核,但是在核膜周围的密度相对较大。 英文
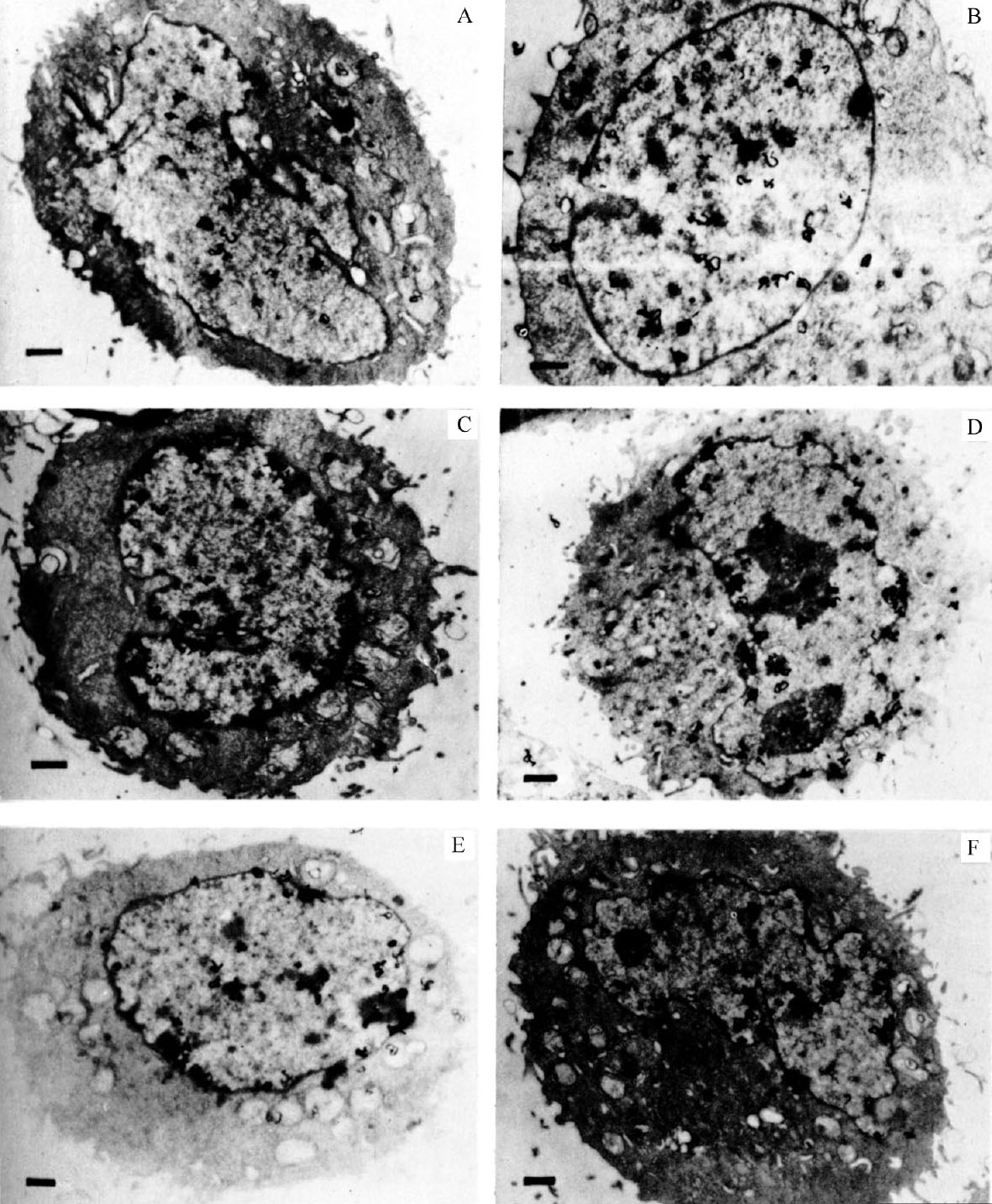
图1. 3 H-TdR处理非同步CHO细胞0.5分钟后放射自显影图。图A和图B展示的是颗粒分布于整个细胞核的情况。图C和图D展示的是颗粒分布在细胞核核膜周围的情况。图E展示的是颗粒的聚集分布情况。图F展示的是颗粒的混合分布情况。用改良的乔克利克基础培养基(购自格兰德岛生物公司)在培养皿中培养CHO细胞,培养基中添加7%的胎牛血清和非必需氨基酸。脉冲标记操作按照以下步骤进行,首先添加5–氟尿嘧啶脱氧核苷(FUDR,购自霍夫曼–罗氏公司)至终浓度为1.6 μg·ml –1 ,以便进一步抑制脱氧胸苷三磷酸dTTP的生物合成。1分钟后,加入 3 H-TdR(51Ci·mmol –1 ,购自新英格兰核公司),至终浓度17 μCi·ml –1 。30秒钟后,将培养皿从37℃中移走,迅速吸掉培养基。然后用含有0.1 μg·ml –1 FUDR的冰浴生理盐水冲洗培养板(换液2次)。室温下,在含有FUDR的生理盐水中进行胰蛋白酶消化,从培养皿中分解细胞。细胞离心后,用戊二醛和四氧化锇固定,脱水,环氧树脂 [29] 包埋,制成紫金色切片。除了用蒸馏水替代醋酸作为定影液,其他的按照卡罗和范·蒂贝根 [20] 的方法将这些切片放在铜网上进行放射自显影。曝光3.5个月。图中标尺代表1 μm。
在一些细胞的研究中发现,许多颗粒距离核膜超过1.25 μm(中间颗粒),这表明复制发生在远离核膜的位点上。但是为了确定这个结论,必须排除其他可能的解释。因为细胞质中几乎没有发现颗粒的存在,所以这些中间颗粒不可能是由于背景造成的。这些颗粒的形成一定是由于放射性标记分子成功结合到DNA分子上,因为在细胞的G1或者G2期也没有发现颗粒的存在(见下文),并且这些从细胞核中分离得到的颗粒可以被DNA酶除去。这些中间颗粒也可能是恰好高于或者低于切片平面的核膜内陷位置上DNA复制的结果。这或许可以解释一些中间颗粒的存在,但绝不能解释大部分中间颗粒的存在。因为这些细胞的切割方向是随机的,我们可以从细胞核外围内陷的频率获悉能让细胞核核膜在这1.25 μm范围内的内陷的几率。尽管图1中有一些明显的周边内陷,但是大部分中间区域与一个内陷之间的距离等于或小于1.25 μm的概率并不高。 英文
我们使用HeLa细胞进行了部分的预实验,在这些实验中,HeLa细胞具有形状更加规则的细胞核,并且在核周围都出现了中间颗粒(图2)。甚至在单细胞0.5 μm厚的连续切片上都有中间颗粒的存在。从S期早期的普遍标记模式到S期后期的周边标记模式的变化说明普遍标记并不是由切割造成的假象。最后,细胞核内部的膜结构并不能解释中间颗粒的问题。其他电子显微镜观察者从未观察到这种膜,除了内陷的周边核膜,我们在切片中也没有观察到这种膜的存在。 英文
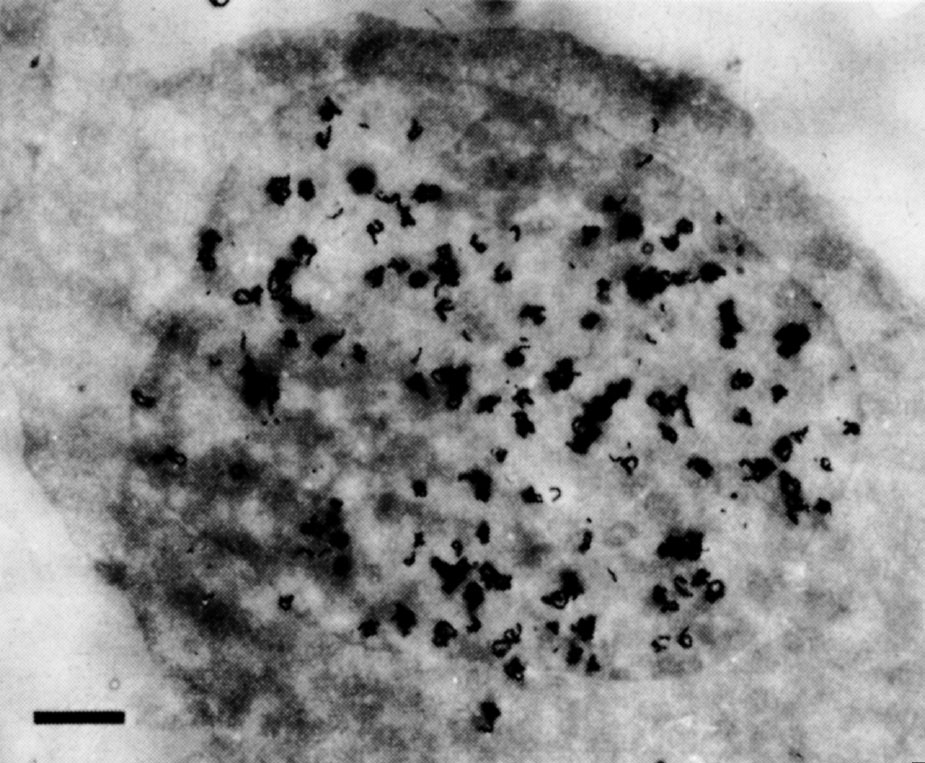
图2. HeLa细胞置于 3 H-TdR1.5分钟后的放射自显影图。用添加5%马血清的改良乔克利克基础培养基(购自格兰德岛生物公司)悬浮培养细胞。加入 3 H-TdR(20Ci·mmol –1 ,购自新英格兰核公司),至终浓度为20 μCi·ml –1 ,进行脉冲标记。1.5分钟后,加入等体积冰浴的含1%戊二醛的生理盐水终止脉冲反应。随后的步骤同图1。曝光9.5个月。图中标尺代表1 μm。
为了定量标定周边标记细胞的比例,我们对威廉姆斯和奥克伊 [16] 的方法进行了改进。我们将那些与最邻近的细胞核核膜相距大于1.25 μm的颗粒定义为中间颗粒,与核膜相距小于1.25 μm的颗粒定义为周边颗粒。一个细胞切片的“中间活性”是以中间颗粒的比例/中间核面积的比例来计算的。这需要排除核仁区域和该区域的颗粒,因为核仁的DNA含量比细胞核其他区域要少。图3显示了非同步细胞群中不同中间活性细胞的数量柱形图(中间活性1.0表示颗粒在中间区域和周边区域分布的密度相同)。 英文
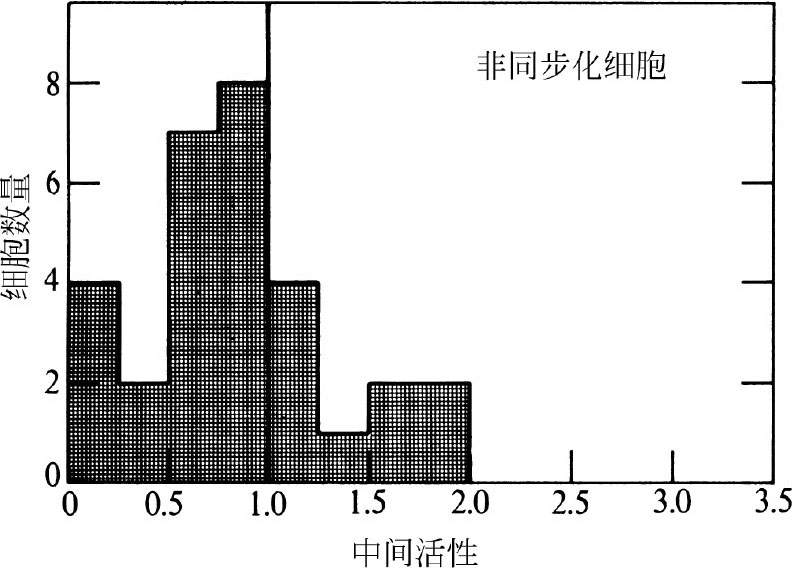
图3. 非同步CHO细胞置于 3 H-TdR0.5分钟后颗粒的分布图。随机选择类似图1和图1中的单个细胞,将其进行放射自显影后以15,000倍放大并打印成图。将细胞核内所有与最近邻细胞核膜相距1.25 μm的点用线连接起来。这条线将细胞核分成中间区域和周边区域。计算每个区域点的数量,并且测量区域面积的大小。在这种计算中,忽略靠近核仁和核仁区域的颗粒,因为核仁区的DNA含量比细胞核其他区域的少得多。位于细胞核外面的颗粒也忽略不计。“中间活性”是计算中间颗粒数量与中间区域面积的比值。该柱形图是对30个细胞核进行测定得到的结果。
图3中仅有13%的细胞中间活性小于0.25,表明DNA合成可以在远离核膜的位点进行。因此在中国仓鼠细胞中,核膜并不是DNA复制必需的。但是许多细胞表现出的显著的周边模式仍然需要进一步的研究解释。 英文
由于前面的放射自显影研究 [14-17] 表明周边标记仅发生在S期的特定时期,我们决定用短脉冲标记处于细胞周期不同时期的同步化细胞。我们采用“秋水仙酰胺” [21] 将细胞同步化。经秋水仙酰胺处理几个小时以后,生长细胞都停滞在有丝分裂期,然后用选择性胰蛋白酶消化收集这些细胞。这些细胞(90%~100%处于有丝分裂期)在缺乏秋水仙酰胺的条件下可以继续生长。2小时之内,98.5%细胞会完成分裂,在完成有丝分裂18小时后,70%以上的细胞会再次分裂。之所以采用这种方法进行同步化而不使用限制核苷酸供应的方法 [21] ,是因为这样不会影响细胞的复苏和正常的DNA复制。 英文
表1和图4显示的是有丝分裂后细胞核内颗粒随时间变化的关系。虽然在S期早期观察不到周边标记,但在S期后期则是以周边标记为主。 英文
表1. 同步化的CHO细胞的DNA合成情况
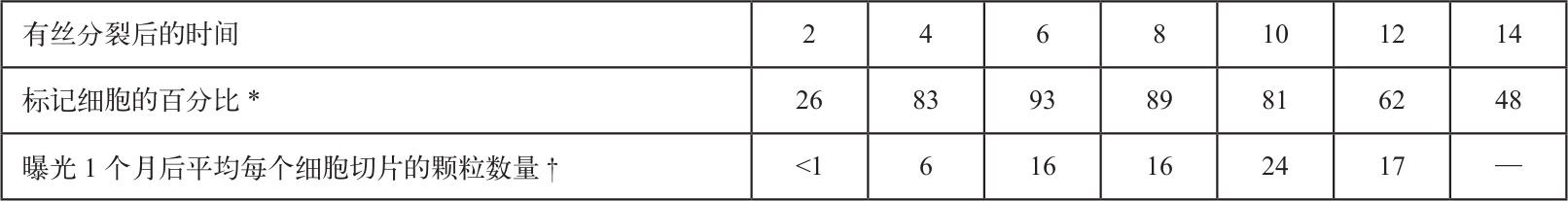
* 细胞的同步化与图4相同。有丝分裂后每间隔2小时,在每个培养皿中加入 3 H-TdR(20Ci·mmol –1 ,购自新英格兰核公司),至终浓度17 μCi·ml –1 。10分钟后,用冷生理盐水洗细胞两次,室温下用胰蛋白酶消化并收集细胞,在低渗溶液(2 mM MgCl 2 ,1 mM EDTA,10 mM KPO 4 ,pH 7.7)中溶胀10分钟,然后收集沉淀,用甲醇–乙酸溶液(3∶1)固定,平铺在玻璃片上风干。用放射自显影乳胶片(“柯达AR-10”)包裹这些玻片,并曝光7天。接着,用吉姆萨氏色素染料染色,然后用“Permount”中性树胶封埋。每一个时间点至少要观察312个细胞来确定被标记细胞的百分比。
† 对图4中用到的放射自显影图片进行了每个细胞核切片上的颗粒的总数的确定。这些图片按照曝光时间进行分类。有丝分裂后2小时和8小时的数据也包括在内。
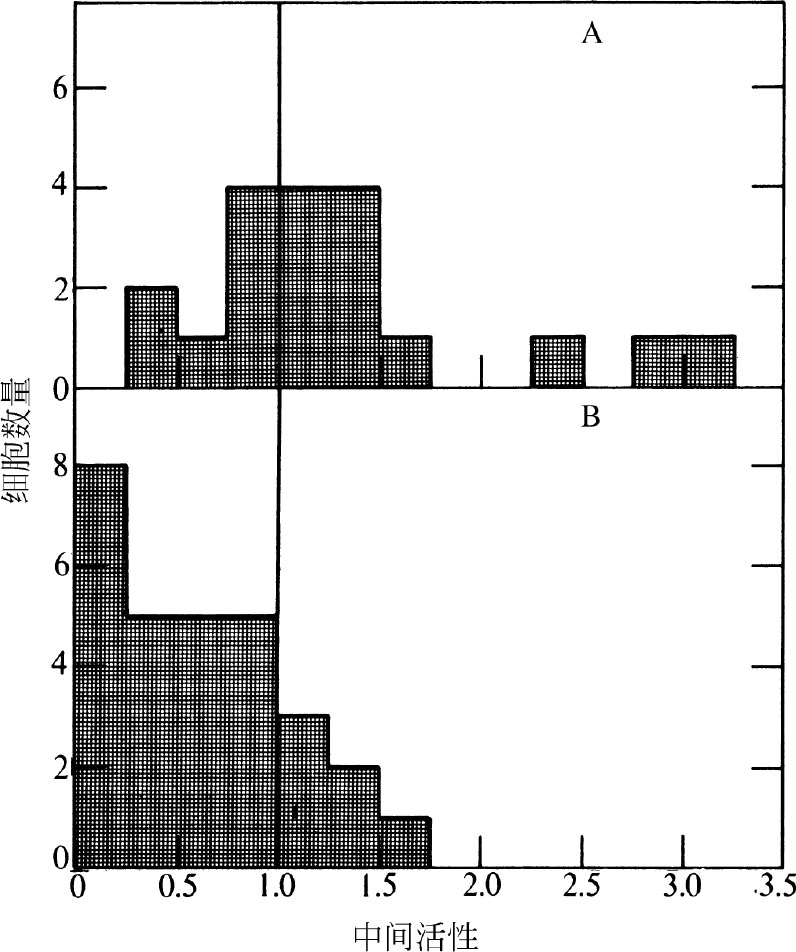
图4. 置于 3 H-TdR下0.5分钟后的同步化CHO细胞中颗粒的分布图。A,S期早期(有丝分裂后4~6小时)。B,S期后期(有丝分裂后10~12小时)。按照斯塔布菲尔德秋水仙酰胺逆转法将CHO细胞同步化。脉冲标记(有丝分裂后每隔2小时进行),电镜切片的准备和放射自显影按照图1的操作。曝光2~4个月。按照图3的方法制作柱形图。A图,有丝分裂后4小时统计了8个标记的细胞核,有丝分裂后6小时统计了11个标记的细胞核。B图,有丝分裂后10小时统计了13个标记的细胞核,有丝分裂后12小时统计了16个标记的细胞核。
如果DNA是在核膜处(或者在细胞核内其他的位置)合成的,那么DNA应该首先转移至核膜进行复制,然后再远离核膜。因而长脉冲或者脉冲追踪形成的颗粒模型应该与短脉冲处理所形成的颗粒模型不同。另一方面,如果DNA复制可以在细胞核的任意区域进行,那么DNA没有必要转移,而且脉冲追踪标记后的颗粒模型应该与简单脉冲标记的颗粒模型一致。 英文
为了验证这些可能性,我们首先脉冲标记处于S期的前半期(有丝分裂完成后的6小时)的同步化细胞0.5分钟,然后用非放射性的胸腺核苷追踪至S期的后期(有丝分裂完成后的12小时)。尽管对这些细胞的检测是在S期后期进行的,但是这些细胞周边标记的比例未如预期,并不比处于S期前半期的细胞高(图4和图5对比)。此外,研究表明脉冲标记非同步化细胞0.5分钟与10分钟,或者先脉冲标记0.5分钟后再追踪6.75小时(图5),其周边标记和普遍标记的比例是相同的。两次追踪实验的结果都显示出有更多的细胞在示踪后被标记,这可能是从细胞内腔持续吸收结合 3 H-TdR的结果。 英文
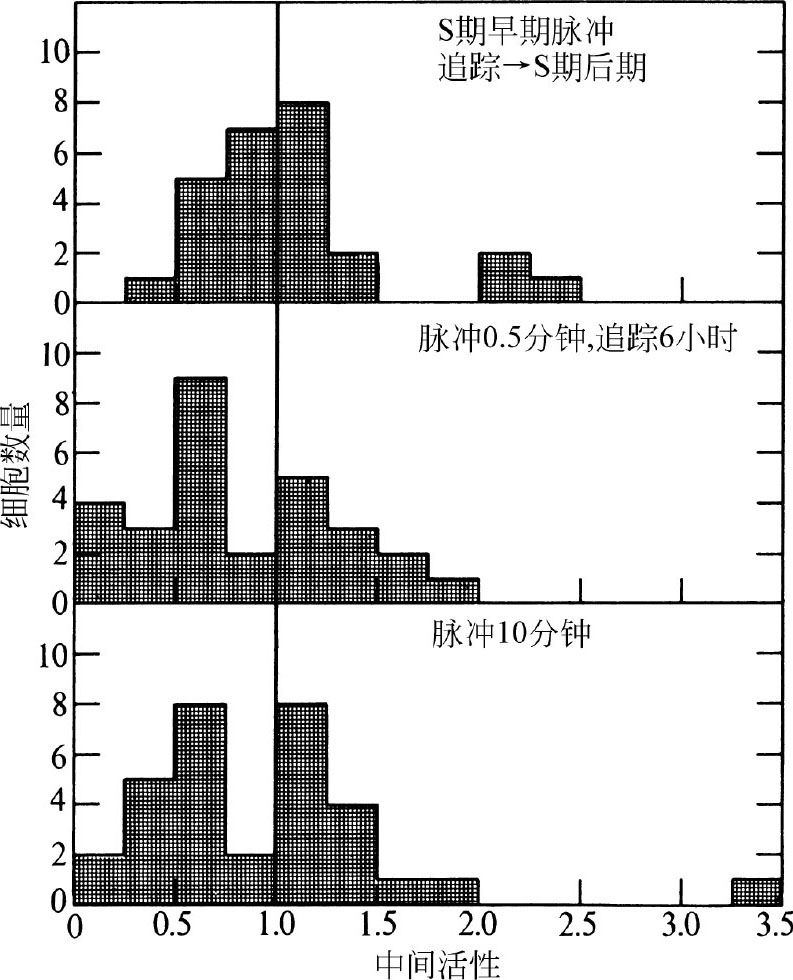
图5. 3 H-TdR脉冲追踪或者长脉冲标记CHO细胞后的颗粒分布图。A,有丝分裂后6小时脉冲标记同步化的细胞(同步方法同图4,标记方法同图1)。标记30秒后,吸掉培养基,冲洗培养皿,然后加入含2.5 μg·ml –1 TdR的培养基。6小时后用胰蛋白酶消化收集细胞,按照图1的方法准备电子显微放射自显影。曝光时间为1.5~4个月。B,按照图1的方法脉冲标记非同步化的细胞,并且同图A一样追踪6.75小时。曝光时间为3.25个月。C,除了用 3 H-TdR处理细胞的时间延长为10分钟,而非30秒外,其他操作同图1。曝光时间为1.5~6个月。按照图3的方法制作柱形图1。图A,统计了24个细胞;图B,统计了29个细胞;图C,统计了32个细胞。
因此,DNA在细胞核内有稳定的定位。由于一些非同步化细胞在追踪过程中已经发生分裂,所以我们的结果还揭示了与核膜紧密结合的亲代DNA的子代DNA也是与核膜紧密结合的。 英文
我们的检测结果显示,0.5分钟和10分钟的脉冲标记模型没有任何差别,所以我们的实验结果可以直接与其他使用长脉冲时间的研究者的结果 [15-17] 进行比较。我们的结果与大多数早期发表的结果一致;普遍标记模型出现在S期早期,周边标记模型则出现在S期后期 [15-17] ,追踪实验还表明,一旦复制开始,DNA在细胞核的位置是稳定的 [15,16] 。 英文
在追踪实验中增加的标记,以及在S期早期明显较低的DNA合成速率(表1),可以解释奥布赖恩等人的放射自显影实验结果 [13] 。在他们的脉冲实验中,S期早期标记的细胞通常会由于每个细胞核中颗粒过少(分布过散)而被忽略;而在追踪过程中增加的标记可以提供足够明显的标记的细胞便于统计。 英文
我们的结果也与科明斯和挂札 [14] 的结果有些不同。他们的报道指出,同步至S期初期的细胞只表现周边标记。威廉姆斯和奥克伊 [16,22] 认为科明斯和挂札的结果可能是由于为了同步化,而将细胞处于长期缺乏核苷酸(用过量的胸腺核苷酸处理24小时,然后用氨甲蝶呤处理14小时)的状态,导致细胞损伤。事实上,科明斯和挂札的实验中所用的同步化细胞的细胞质含有许多大的空囊泡(这表明细胞严重损伤),而他们实验中的非同步化细胞的细胞质却正常。细胞损伤导致周边标记模式的原因尚不清楚。这或许与目前发现的由于多瘤病毒感染而导致的鼠卫星DNA复制早于主体DNA复制 [12] 的情况有些关联,通常情况下卫星DNA的复制会相对较晚 [23-25] 。 英文
由于出现细胞损伤,科明斯和挂札 [14] 结果的可信度大打折扣,他们的结果让人误以为DNA的合成发生在整个细胞核区域,且核膜没有参与复制的起始。然而我们的研究发现,在S期较早期(有丝分裂后2、4小时),每个细胞的平均复制速率(细胞核经过固定的曝光时间标记以后,每个核切片的平均颗粒数)要远低于S期后期(表1)。这表明,在真正的S期初期,复制速率太低以至于检测不到标记,更无从确定是周边模式还是普遍模式。 英文
已经证明异染色质的复制比常染色质晚,在分裂间期以浓缩的状态存在 [26-27] 。我们以及其他研究者的切片观察 [14-17] 表明,浓缩的染色质主要与内核膜相连。因此我们的结果与以下的想法 [16] 一致:常染色质的复制发生在S期早期而且分布在细胞核的整个区域,而异染色质的复制发生在S期后期,在内核模和其他分散的区域以浓缩的状态存在。 英文
尽管许多关于DNA在细胞核内合成位点的细胞分级分离实验 [8-11,13] 已经被诠释为倾向于支持复制位点与细胞核核膜相连的观点,但是这并不是唯一可能的解释。同样合理的一种可能性是,细胞裂解后,由于生长点的某种特殊结构属性(如大多是单链),可能导致其与体内其他大部分的DNA相比能够结合更多的蛋白、膜或者其他物质。当然,也可能生长点确实与细胞内的一些物质相连,造成生长点与体内主体DNA在细胞裂解分离过程中分离。 英文
因为在哺乳动物细胞 [8-11,13] 和细菌细胞 [2,3,5] 上开展的实验研究使用了相似的实验技术,并得到相同的结果,所以在细菌体内生长点很有可能也不与膜相连。 英文
特别感谢伊莱恩和罗伯特·伦克提供的建议和帮助。该项目受国家科学基金会和美国国立卫生研究院的资助。电子显微镜实验在麻省理工学院生物系电子显微镜实验室进行。 英文
附加说明: 最近福孔等人 [28] 用小鼠细胞进行自显影和细胞分级分离实验,结果与我们的结果类似。 英文
(吕静 翻译;刘京国 审稿)