
下载掌阅APP,畅读海量书库
立即打开



 Eukaryotes-Prokaryotes Divergence Estimated by 5S Ribosomal RNA Sequences
Eukaryotes-Prokaryotes Divergence Estimated by 5S Ribosomal RNA Sequences
M. Kimura and T. Ohta
Editor’s Note
This brave stab at estimating one of the earliest but most crucial events in evolution marks an early attempt at using comparative molecular data to calibrate evolutionary history. Motoo Kimura and Tomoko Ohta compared sequences from 5S ribosomal RNA—one of the most “ conserved ” of all genetic sequences, meaning that it differs little across species—to estimate the divergence date between eukaryotes (such as humans and yeast) and prokaryotes (here two bacterial species). The estimate of two billion years would not surprise anyone today, but it caused some surprise then by implying that yeast is more closely related to humans than to bacteria. 中文
DATING the principal events in the history of life on the Earth is an interesting subject in evolutionary studies. Here we estimate the time of divergence of the eukaryotes and the prokaryotes through comparative studies of 5S ribosomal RNA sequences, coupled with those of cytochrome c . By prokaryotes we mean primitive forms having no true nucleus (bacteria and blue-green algae), while by eukaryotes we mean higher nucleated organisms such as plants and animals including yeasts and fungi. The principle we use in our estimation is that the rate of nucleotide substitutions in the course of evolution is constant per year per site for each informational macromolecule as long as the structure and function of the molecule remain unaltered. 中文
To estimate the evolutionary distances (number of mutant substitutions) among 5S rRNA sequences, we made the alignment shown in Fig.1, using published data 1–3 on human, yeast and bacterial ( Escherichia coli and Pseudomonas fluorescens ) sequences. To arrive at this alignment, previous attempts 1,3 involving two or three sequences were helpful. The alignment is made in such a way that the number of matches between sequences is maximized while keeping the gaps inserted as few as possible. It involves a trial and error process, shifting various regions, and counting the number of nucleotides by which the two sequences agree with each other, followed by calculation of probability that this or better agreement occurs by chance. Figure 1 shows clearly (as was noted already by others) the marked conservative nature of this molecule as shown by the fact that only a small number of gaps need be inserted to obtain homology. The observed differences between sequences in terms of the fraction of different sites are given in Table 1. The mutational distance, that is, the average number of nucleotide substitutions per site, was estimated using the formula
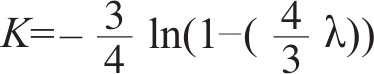
where λ is the fraction of sites by which two homologous sequences differ. The formula was derived under the assumptions that in the course of evolution nucleotide substitutions occur spatially at random and with uniform probabilities and that each of the four bases (A, C, G, U) mutates to any of the remaining three with equal probability. (For details see ref. 4.) The equivalent formula has been previously derived by Jukes and Cantor 5 . We should also note that Dayhoff’s 3 empirical relationship tabulated in her Table 11–3 is practically equivalent to this formula especially for the purpose of comparing different K values. 中文
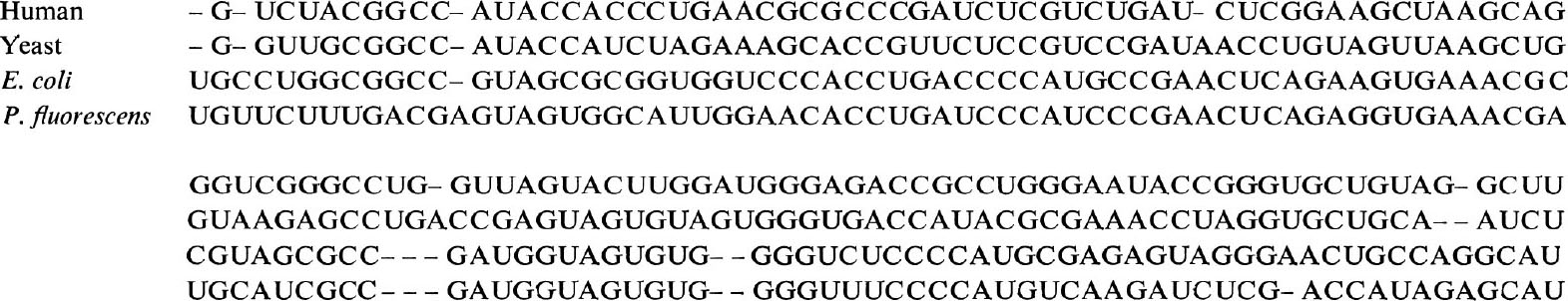
Fig. 1. Alignment of 5S rRNA sequences.
Table 1. Fraction of Different Sites between 5S rRNA Sequences
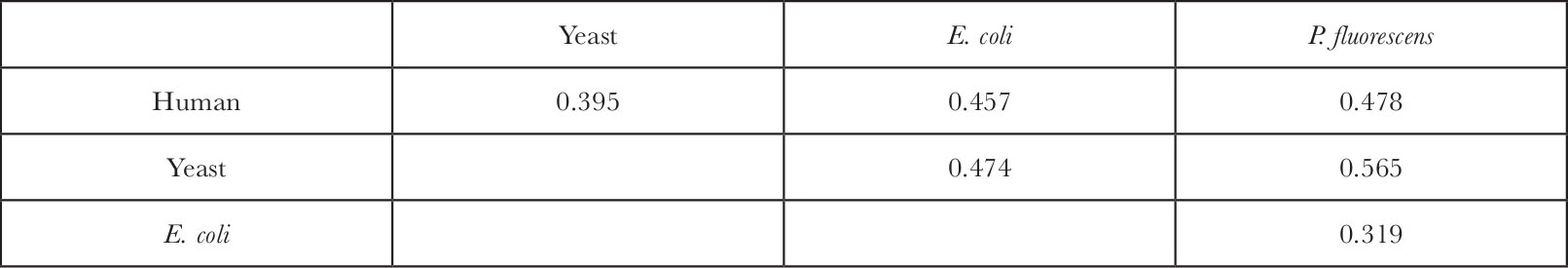
The average mutational distance between the eukaryotes (man, yeast) and the prokaryotes ( E. coli , P. fluorescens ) turned out to be
K eu–pro =0.817±0.158
where the standard error was obtained from four observations (comparisons). On the other hand, the corresponding quantity between the human and yeast was
K h–y =0.561±0.095
where the error is a theoretical one computed by equation (5) in ref. 4. It may be interesting to note here that yeast is more closely related to man than to the bacteria, supporting the thesis (compare ref. 6) that the division between the eukaryotes and prokaryotes is more basic than divisions within eukaryotes. The remoteness of the eukaryotes-prokaryotes divergence relative to the human-yeast divergence can be estimated by the ratio K eu–pro / K h–y which is approximately 1.5. This is much lower than the corresponding estimate of McLaughlin and Dayhoff 7 who obtained the ratio 2.6 using data on cytochrome c , c 2 and tRNA sequences. It is also lower than the corresponding estimate, ~2, obtained by Jukes (1969) (quoted in ref.7). Hoping to resolve this discrepancy we calculated the evolutionary distances using the same data 3 on tRNA as McLaughlin and Dayhoff 7 but restricting our treatment only to paired regions. The reason for doing this is that it seems as if there is no excess of highly conserved regions (as inferred from our statistical analysis of the frequency distribution of the number of evolutionary changes per site in the alignment of Fig. 1), so that evolutionary change appears to be uniform over the entire sequence and in this respect 5S rRNA might be more similar to the paired than unpaired regions of tRNA. The evolutionary distances turned out to be K 1 =0.836±0.136 for the eukaryotes-prokaryotes divergence but K 2 =0.420±0.029 for the average of rat-yeast (tRNA Ser ) and wheat-yeast (tRNA P he ) divergences. It may be seen that although K 1 is comparable to K eu–pro , K 2 is clearly lower than K h–y , so that the ratio K 1 / K 2 =1.99 is still considerably higher than K eu–pro / K h–y =1.46. It is possible that the difference is due to sampling error, with the true value lying somewhere between these two. At any rate we should take these estimates as tentative (including the problem that might arise because of the multiplicity of ribosomal genes). There is some reason to believe, however, that our estimate of 1.5 is consistent with the fossil records as explained below. 中文
From comparative studies of cytochrome c sequences among eukaryotes, we can estimate the remoteness of the human-yeast divergence relative to the mammal-fish divergence. This allows us to estimate the absolute time of the human-yeast divergence as it is known from classical palaeontological studies that the common ancestor of the fish and the mammals goes back to some 400 m.y. (compare refs 8 and 9). From a number of comparisons involving various species of fish and mammals (data taken from ref. 3), we obtained the results that the mutational distance (in terms of the number of amino acid substitutions) between mammals and yeast is about three times that between mammals and the fish. This puts the time of the human-yeast divergence back to about 1.2×10 9 yr. This agrees with Dickerson 10 who obtained 1,200±75 m.y. as the estimated date of the branch point for animals/plants/protists. We should note here that we avoided using the cytochrome c 2 sequence of the bacterium Rhodospirillum rubrum to estimate the prokaryotes-eukaryotes divergence, because there seems to be some difference in function between cytochromes c and c 2 (compare ref. 7). 中文
Multiplying 1.2×10 9 yr by the ratio K eu–pro / K h–y ≈1.5, we arrive at the result that the divergence between the eukaryotes and prokaryotes goes back to some 1.8×10 9 yr. Recent studies on Precambrian fossils (compare ref.11) suggest that the eukaryotes evolved from prokaryotes at some point between the Bitter Springs formation (10 9 yr old) and the Gunflint formation (2×10 9 yr old). With additional relevant data forthcoming (for example, ref. 12) we hope that the studies of molecular evolution will soon supply a more accurate date. 中文
We tentatively conclude that the eukaryotes diverged from prokaryotes nearly 2×10 9 yr ago, thus opening up the way toward “higher organisms”. 中文
We thank Dr. K. Miura and Mr. H. Komiya for calling our attention to the relevant literature on 5S rRNA sequences and Drs. T. Maruyama, S. Takemura and S. Kondo for helpful discussions. 中文
( Nature New Biology , 243 , 199–200; 1973)
Motoo Kimura and Tomoko Ohta
National Institute of Genetics, Shizuoka-ken 411, Mishima
Received December 29, 1972; revised March 5, 1973.
References: