
下载掌阅APP,畅读海量书库
立即打开



 大鼠与小鼠桑葚胚的融合以及嵌合囊胚的形成
大鼠与小鼠桑葚胚的融合以及嵌合囊胚的形成
泽尔马克
编者按
十多年前,波兰胚胎学家安杰伊·塔尔科夫斯基通过融合不同种系的小鼠卵细胞,培育出了嵌合小鼠。在本文中,赫拉德·泽尔马克尝试将小鼠与大鼠的早期胚胎融合得到嵌合囊胚,也就是包含了小鼠细胞和大鼠细胞并且有了些许进一步发育的胚胎结构。这项研究为可育的种内嵌合体的产生,比如山羊和绵羊的杂交体即“山绵羊”,奠定了基础,这帮助人们回答了一些关于发育的基本问题。在人类DNA与牛、兔的卵细胞融合实验中观察到,在不需要人类卵细胞的条件下,早期嵌合体就能产生人类干细胞,这在干细胞研究中跨越了一个大的障碍。这些产生嵌合体的技术还有助于挽救濒临灭绝的物种。 英文
通过将不同种系小鼠的囊胚细胞融合诱导产生嵌合体是近期实验胚胎学领域最卓越的成就之一 [1,2] 。 英文
去年,有人(麦克拉伦博士,私人交流)指出,将不同物种的桑葚胚融合在一起的可能性,这种可能性还没有得到充分的研究 [3] 。因此我用与融合小鼠桑葚胚基本相同的技术 [3,4] 进行了一系列的实验,来研究能否诱导大鼠和小鼠桑葚胚的聚集。 英文
在妊娠第2天的21:00,用培养液将小鼠桑葚胚(C3Hf×瑞士种,8细胞至16细胞阶段)从子宫–输卵管区域洗脱出来,未经历栓塞,见光14小时,置于暗处10小时,并使黑暗阶段的中期处于午夜。在妊娠第3天的21:30,洗脱出大鼠桑葚胚(R,阿姆斯特丹大鼠,8细胞至16细胞阶段)。分离出来的桑葚胚分别储存在37℃下油封的含丙酮酸的培养基中,培养空间空气中的CO 2 含量保持在5% [5] 。 英文
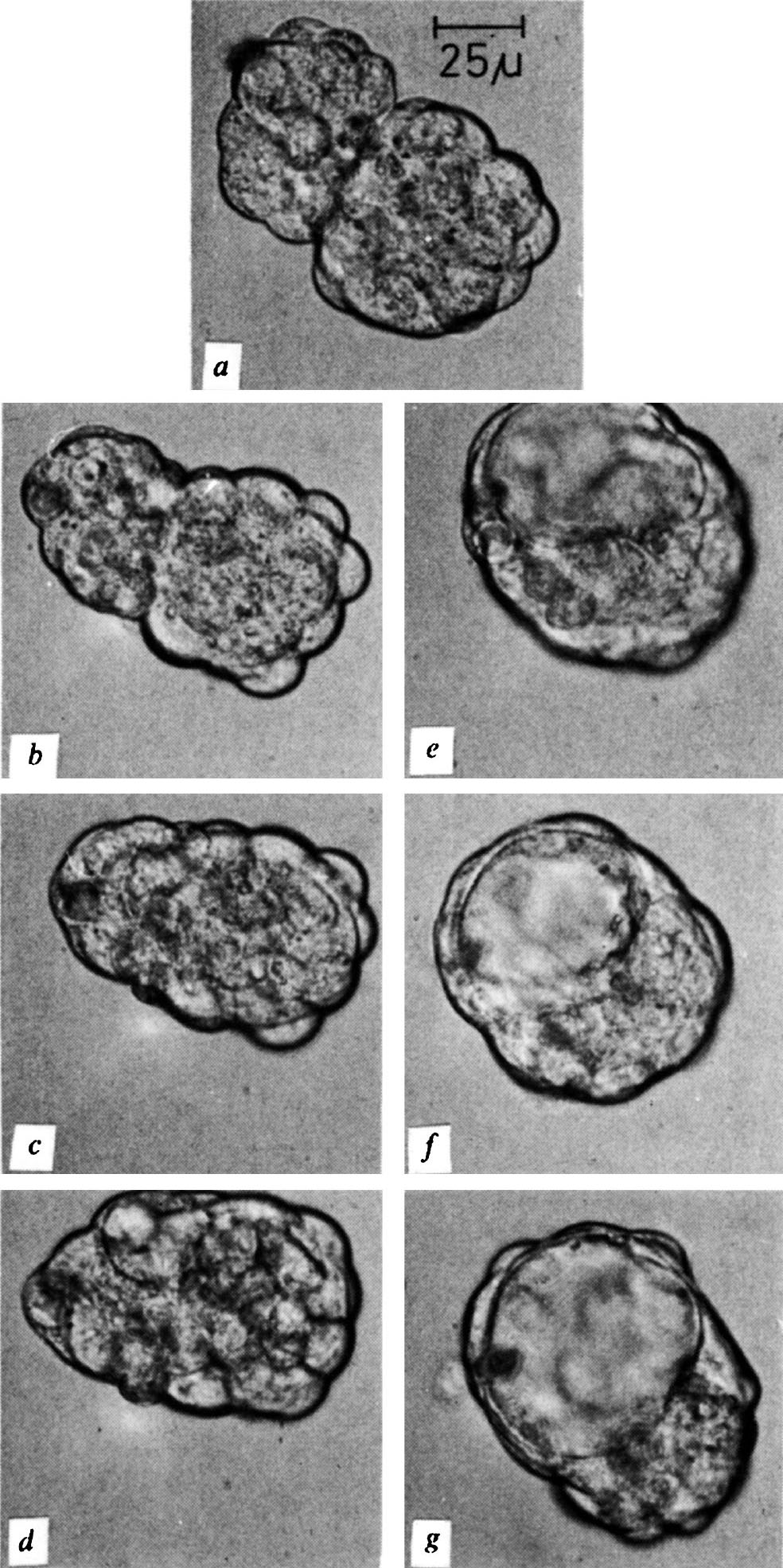
图1. a ,撤掉胚胎固定器后不久,小鼠(左侧)和大鼠的桑葚胚紧邻。图中显示经过2小时( b )、4小时( c )、6小时( d )、8小时( e )、11小时( f )和12.5小时( g )后的发育阶段。在 d 阶段,囊胚腔已经形成。
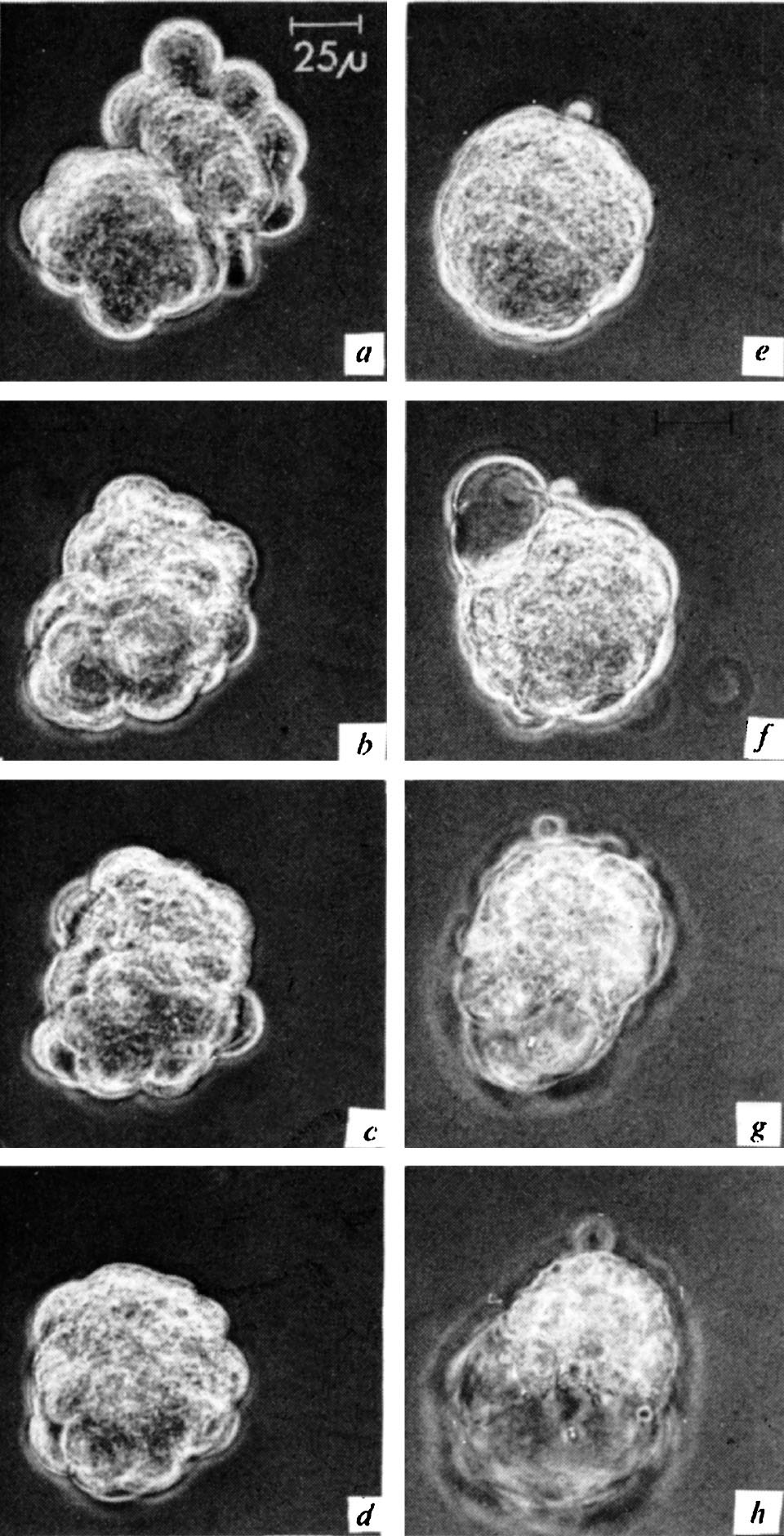
图2. 融合过程中的小鼠(左侧)和大鼠(右上)桑葚胚。图片采自融合开始30分钟( a )、2小时( b )、6小时( c )、8小时( d )、12.5小时( e )、18.3小时( f )、24小时( g )和34小时( h )后。( e )阶段仍可见到大小鼠桑葚胚的界线。第一空腔在( f )阶段形成,最终的空腔在( g )阶段形成。
将卵细胞在0.5%的链霉蛋白酶中温育3到5分钟即可溶解透明带 [6] 。在培养基中,通过压迫卵细胞穿过狭窄的移液管从而最终去除透明带。在每个实验中,借助两个由显微操纵器驱动的含有封闭腔 [7] 的卵细胞固定器,一个小鼠的桑葚胚和一个大鼠的桑葚胚在37℃下的油滴培养中彼此接触。在这个持续20到30分钟的接触阶段,培养皿并不关闭,使潮湿气流直接吹向油表面以维持培养基的pH和渗透压。 英文
移除卵细胞固定器以后,用连接在倒置显微镜上的间隔定时摄像机持续追踪这一对桑葚期胚胎的发育过程,每隔30秒打开闪光灯进行一次照相。 英文
一共进行了16组相同的实验,其中有12组发生了成功的胚胎聚集性融合。根据在巨大的桑葚胚表面正在分裂的细胞的暂时性突起可以看出,融合双方都进行着活跃的增殖。有时通过细微的压痕可以鉴定出融合很长时间后的单个桑葚胚。囊胚腔形成的方式是多样化的。在某些情况中,它出现在能够在较早期看到的两个桑葚胚的连接位置(图1 b ~ d );而在其他情况中,发育不完全的囊胚腔是在其中一个胚胎中形成的(图2 f ),并在之后消失。在同一个融合胚胎中,最终的囊胚腔在另一个参与融合的胚胎内部形成(图2 g )。在一组实验中,参与融合的胚胎之一在细胞团变圆之前就形成了腔(图3 b ),然后消失,最终的腔是在融合之后形成的(图3 d )。间隔定时拍摄的图片和分析显示大鼠和小鼠细胞均参与了巨大囊胚的形成。 英文
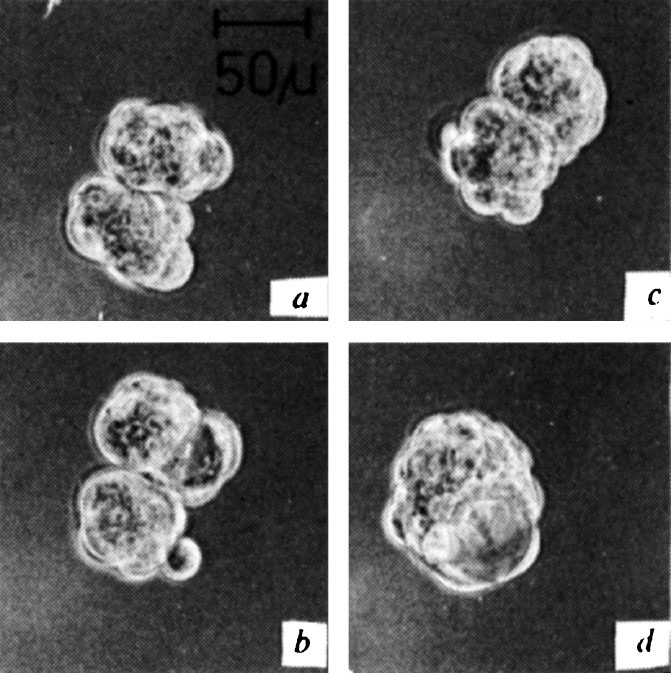
图3. 在小鼠桑葚胚内形成腔并消失以后,大鼠和小鼠的桑葚胚发生融合。图片拍摄于融合开始大约30分钟( a )、6.6小时( b )、10小时( c )和19小时( d )。
嵌合囊胚的总体特征就是内细胞团相对较大。 英文
初步的观察显示大鼠卵细胞的发育阶段对成功的融合至关重要,而且使用显微操作器非常有帮助。 英文
我们可提供间隔定时拍摄所获得的录像拷贝用于研究。 英文
(毛晨晖 翻译;梁前进 审稿)