
下载掌阅APP,畅读海量书库
立即打开



 任何一个地层中众多中间变种的缺乏
任何一个地层中众多中间变种的缺乏
通过前面的探讨,我已经得出结论,地层对古生物的记录无疑是非常不完整的。如果我们仅单一地去研究某一类地层,那么便无法理解,为什么在这个区域生存 的有亲缘关系的物种,长久以来稳定地生活在这里,但是却找不到与它们关系紧密的中间类型。
有人曾对同一块地层的上下层物种进行对比,发现了清晰的变种关系。
比如,特劳希勒对
菊石
 的记载;又如,黑尔甘道夫勘察瑞士的淡水沉积层时,发现其中多个地层 中都有多形扁卷螺,它们奇异地呈现出10种递变类型。
的记载;又如,黑尔甘道夫勘察瑞士的淡水沉积层时,发现其中多个地层 中都有多形扁卷螺,它们奇异地呈现出10种递变类型。

菊石
除此之外的记录就非常稀少了。沉积层的形成无疑需要经过漫长的岁月,对于始终生活在某个区域的物种来讲,地层的沉积作用为什么不能将它们之间的递变演 化记录下来呢?对下面的一些解释,就目前的研究水平来看,我还不能做出恰当的评价。
关于地层形成时间与物种演变的探讨
地层的形成,相对于物种演变,要更加漫长一些。古生物学家勃龙和伍德沃德认为,地层的平均形成时间是物种的平均
续存
时间的两三倍。他们的观点是非常值得尊重的,但我认为,现实还存在着许多无法克服的困难,让我对此难以做出恰当的评论。
当我们在某个地层中首次发现一个物种时,就断定为新生物种,否认它以前可能在其他地方出现过,这未免过于轻率。同样,当我们发现某个物种在地层的形成 结束前就消失了,就判断它为灭绝,这个结论也不够全面。欧洲的面积比起整个地球的其他区域,是何其之小,这一点我们不该忘记,而且整个欧洲相同地层的各个阶 段也不是完全对应的。
由于气候的改变,所有海生生物都曾进行过大规模的迁徙,这一点我可以谨慎地给出结论。当我们在一个地层首次发现某种生物时,只能断定这个生物刚刚迁徙 到这个区域。众所周知,很多在北美洲和欧洲都出现过的生物,在北美洲古代地层出现的时间要早于在欧洲地层出现的时间。显然,它们从北美洲迁徙到欧洲需要很长 的时间。
我们考察现代沉积物时,经常可以在沉积岩内找到现存物种,但是在周围的海域中却很难找到这些物种;与之相反的是,在周围海域内广泛生存着的物种,却很 难在附近的沉积岩中被发现。这种现象在地球上比比皆是。
对
冰河时期
(地质时期的一小部分)的一些数据进行研究,包括欧洲生物的迁徙量、海陆的升降 情况、极端气候的变化、时间的历程,都是非常有意义的。世界各地含有化石的沉积层,是否在整个冰河时期,都能在同一个地区连续不断地进行沉积,对此我表示怀 疑。
密西西比河
河口附近生存着数量繁多的海生生物,但在冰河时期这些物种形成的沉积物都能按照 时间顺序一层层地积累起来吗?我们清楚地知道,在同一历史时期,美洲的其他地区曾发生过巨大的地理变迁。冰河时期的早期,在密西西比河河口附近形成的沉积层 ,经历过上升期后,地理位置和生存在这里的物种均发生了改变。这时,早前的沉积物有可能被侵蚀,而新的物种有可能被掩埋,并首次出现在沉积层中。未来,某个 地质学家来勘察这块地层时,有可能会做出错误的判断。地层中发现的生物,平均生存时间是短于冰河时期的,然而全面地分析这些生物遗骸的沉积情况后,会发现有 些生物的出现时间是早于冰河时期的,并且一直延续生存到今天。
只有沉积层保持长时间不断的稳定积累,时间长到足够使物种完成缓慢的变异过程,产生新的物种时,同一地层的上下部分才有机会记录下两个物种之间出现的 中间类型。这个过程产生的地层一定是非常厚的,而变异的物种也必须始终生活在这个地区。
通过前面的探讨,我们已经了解到, 地层稳定沉积的条件,一定是在地层下沉后,沉积物的供给等同于下沉量,海水的深浅没有发生太大变化,生物体能 稳定地在这里继续生存。 但是问题来了,随着下沉运动的产生,原本供给沉积物的地区也会下沉到海中,当多次连续下沉以后,沉积物的供给将会越来越少。 所以,沉积物的供给量很难等同于下沉量,于是很多古生物学家发现,在一块深厚的沉积层里,除了靠近地层顶部和底部的区域以外,其他区域都很少会发现生物化石 。
地层与物种定义
地球上每个单独的地层,和其他任何地方的地层一样,沉积作用往往被间断过。我们看到一个地层是由完全不同的矿物质组成时,更能说明这个地层的沉积作用 曾被中断过,而且这种现象是很常见的。无论我们多么仔细地去研究一个地层,都很难搞清楚它的沉积时间。
在很多例子中我们都发现,有时一块几英尺厚的岩层沉积时间,等同于其他区域一块上千英尺厚的岩层形成时间。如果你忽视了这个问题,那么对于某些地区薄 薄的地层,你可能很难与漫长的岁月联系起来。
以下这种情况也很常见:
一个地层升起以后,被侵蚀,然后再次下沉,在它的最上层又堆积了与前面相同的沉积物,中间的沉积间断是非常容易被人们忽视的。
更为奇特的是:
某些巨大的树木化石依然保持活着时候的直立状态。
硅化木,一种常见的木化石。

这也说明,在沉积过程中,由于水平面的变化和沉积间断期的存在,才使这些树碰巧保存下来。
莱伊尔爵士和道森博士曾在新斯科舍省发现了1400英尺厚的石炭纪地层,其中含有古代树根的化石。它们一层一层地叠起来,形成不少于68个不同的水平面。
如果在某个地层的上、中、下三个部位,都发现了同一个物种,那么这个物种在整个沉积期内并没有生存在同一个地点,而且在同一地质时期出现了几次灭绝和 重现。
同时,假如某个物种发生了明显的变异,理论上应该存在的,与它有着细微变化的中间类型,是不会在地层中被找到的。
物种和变种
博物学家对物种和变种并没有特别严格的定义。他们认为,物种之间或多或少都存在差异,当两个类型具有较大差异,且找不到它们之间的中间类型时,便将两 种类型各自定义为一个物种。
然而,前面我们曾讨论过,在任何一块地层中,不同的断面之间很难发现我们期望的过渡关系。
假设A是存在于地层下层的古老生物类型,B和C是另外两个物种。
倘若A实际为B和C的中间型,这时如果我们找不到A分别与B和C之间紧密的关系时,我们就会将A单独定义为一个物种。
文章的开始我也曾讲过,倘若A是B和C的祖先,它也不会完全表现出两者中间型的特征。所以,很可能我们在地层的不同层面发现了亲代和多种变异后的子代, 然而它们的中间过渡型却消失了。这时,我们找不到它们之间的亲缘关系,于是将它们分别定义为不同的物种。
很多古生物学家在鉴定生物化石时,仅仅根据它们之间非常细微的差异就将其各自定义为一个物种,尤其当这些标本不是取自同一地层时,更是如此。这么做未 免考虑不够全面。
被多比内和其他学者区分过细的许多物种,在一些经验丰富的贝类学家那里已经被降级为变种。这一论证也为物种演变提供了理论基础。多数博物学家认为,发 现于第三纪末期沉积层中的许多贝类,与现存的物种是一样的。而著名的博物学家阿加西斯和皮克特却认为,第三纪的物种尽管与现存物种差异并不大,但也应该划分 为独立的物种。
在这种情况下,如果我们赞同以上两位博物学家的论证,认为第三纪的物种确实不同于现代物种,而与大多数博物学家的意见相左时,却恰恰找到了物种一直在 进行频繁的细微变异的证据。
我们再来对一块地域广阔的地层进行观察,同时我们将观察间隔拉长,那些相互连接但层位不同的地层中掩埋的化石,虽然看起来是不同的类型,但拿来与相隔 更远的地层中的化石相比,同一块地层中化石的亲缘性可就近多了。不难看出,我们又发现了验证物种演变理论的证据,具体内容我们后面再说。
结合前面讨论过的内容,我们可以推断: 繁殖量大、生活区域较固定的动植物,最初的变异仅出现在局部地区,当这些变异完成了某种程度上的改变时, 它们才有可能扩大自己的生存空间,排挤掉原来的物种。 这种从最初变异到完成很大程度上的改变并成为新类型,往往都是局限于某个区域的,所以想要在任 意一个地层中找到两个物种之间的全部过渡类型,概率是非常小的。
就植物来讲,往往分布范围较广的物种,最容易出现变异。大部分海生生物的分布范围是非常广的,尤其是贝类和那些散布到欧洲地区以外的海生生物,它们更 容易出现地方变异,最后演化为新的物种。它们的生存范围远远超出欧洲地层的界线,所以我们想在某块地层中找到物种演化的一系列过渡阶段,其机会便大大降低了 。
福克纳博士最近发表的新观点,与我的看法相同。那就是,每个物种演化的时间,用年代来计算的话是很长的,但与物种没有任何变化的时期相比,还是短暂的 。
我们应该了解,直到现在,能用化石将两个物种的中间类型连接起来的情况是非常稀少的。我们即便从很多地方采集大量的化石,都很难证明两个物种之间的关 系。而且,采集化石本身就是一件比较困难的事。
生活在北美洲岸边的某些贝类,它们的祖先是同一个物种,还是几个不同的物种呢?
一些贝类学者认为它们与欧洲有代表性的贝类不同,将它们分别定义为一个物种;还有些贝类学者认为它们只是变种。
只有在获得大量的生物化石,搞清它们之间全部的过渡类型才能得到答案,然而这几乎是不可能完成的事。
那些认为物种不会演变的学者不断地强调,地质学上找不到物种之间的过渡类型,这个说法我会在下一章进行详细反驳。
正如卢伯克爵士所说的,“每一个物种都是其他近似物种的中间过渡型”。假设某个属一共有20个种(包括现存种和灭绝种),其中4/5已经灭绝,那么现存种 之间的差异看起来就会比较大。再假设这个属中的两个极端类型也不幸灭绝了,那么这个属与近似属看起来区别就更大了。
我们无法做到的事,却不断地被提出来,同时作为反对我的理论的有力证据。
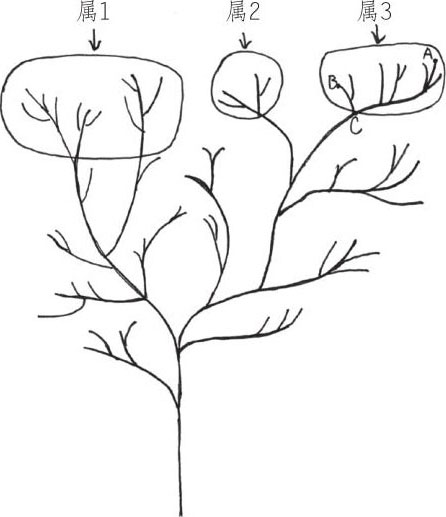
属1属2属3以属3为例,如果该属中只有A、B和C这3个物种还存在,其余物种均灭绝,那么A和B的差异性从图中就可以看出是比较大的。如果A和C也灭绝了,只剩下B,那么B代表的物种和属1、属2中物种的差异就更大了。但是,这3个属都是由一个共同祖先演化而来的。
不幸的是,地质学的研究结果都没有显示,过去曾出现过无数中间类型,如同现代的变异一样,它们之间存在着细微的差别,但能将所有物种——无论是现存物 种还是灭绝物种——联系起来。这是
关于马来群岛的假想
以上造成地质记录不完整的原因,我用一个假想的例子将它们概括起来。
马来群岛的面积与欧洲面积(从北角到地中海,从英国到俄罗斯)差不多,与除美国外,已经测量过的地层面积的总和也差不多。我与戈德温·奥斯汀先生的观 点相同,马来群岛的无数个岛屿散落在广阔的浅海海域,这与处在地层沉积时期的远古欧洲情况相似。栖息在马来群岛的生物种类繁多,这里是世界上物种最丰富的地 区之一。但倘若把曾经在这里生活过的物种全部搜集起来,就会知道这个世界自然史代表是多么不完整!
我们有足够的理由相信,在假设的这个马来群岛上,多种情况下生物遗骸不能被保存下来。首先,陆生生物肯定难以被完整地掩埋而形成化石。在岸边栖息生活 的动物,在海底裸露岩石上生存的底栖动物,能被掩埋的概率也不高。就算它们被砾石和沙子盖住了,也很难被长久地保存下来。海底是否有沉积物不断产生,或者沉 积物的沉积速度是否够快,决定了生物遗骸在腐烂之前是否能够被掩埋。
马来群岛若要形成与中生代相似的深厚地层,并且其中还蕴含丰富的化石资源,只有地层发生沉陷才有可能实现。当某块区域下沉后,其他区域可能保持静止或 上升,在各个区域之间便产生了长久的时间间隔。在上升时期,靠近陡峭海岸的地方,已经形成的化石层,在不断堆积的同时又不停地被波浪侵蚀着,堆积与侵蚀作用 几乎等速,如同我们在南美洲海岸看到的那样。
同时,地层上升后,即使广阔的群岛都处在浅海中,后来的沉积物也无法掩盖之前的沉积层。沉积层不能继续增厚,就不可能保存到久远的未来。下沉时期,容 易发生物种灭绝;上升时期,容易出现物种变异。无论如何,地层都无法完整地对此进行记录。
全部群岛的下沉或者部分群岛下沉经历的时间,即沉积物的堆积时间,与物种的平均生存时间之间的关系,更是难以解释清楚。两个或多个物种之间出现的所有 过渡类型能否保存下来的一个非常重要的条件,便是地层的沉积速度与物种生存时间的同步程度。
如果其中的过渡类型不能被完整地保存下来,那么仅存的生物类型将会被定义为亲缘性非常近的新物种。在下沉时期的漫长岁月中,沉积作用可能会因水平面的 震动而打断。
同时,气候的细微变化,也会使生物的栖息地发生变化,部分生物会移居到其他地方。这样看来:
在任何一块地层中都很难保存生物变异的全部证据。
原本生活在群岛的大部分海生生物,现在已经离开群岛,生存于远在千里之外的地方。不难推断,这些分布广泛的物种,更容易发生变异,产生新的变种,虽然 发生变化的只是它们中的一部分。
最开始,变异的出现是有局限性的,可能仅仅发生在某一地区,当它们具备某种决定性的优势后,也可能是变异得到进一步改良,它们会逐渐扩大,并取代原来 的亲种。
当这些变种重新回到原来亲种生活的区域时,由于它们与亲种之间已经出现了差异,虽然差异可能并不大,但当它们与亲种同时在地层的不同亚层中被发现时, 按照许多古生物学家的分类原则,它们将会被定义为新物种。
如果前面的假设成立,那么我们就不可能在地层中发现无数中间类型。依据我的学说来看,中间类型能将所有现存的和灭绝的物种关联起来,最终编织成一个非 常长的、具有无数条分支的生物遗传链。当我们寻找一小部分遗传链时,确实发现了一些连接关系,其中的生物有的亲缘关系较近,有些却非常远。而处在连接关系中 的生物型,就算亲缘关系非常密切,当它们在地质层不同的层位被发现时,还是会被定义为不同物种。
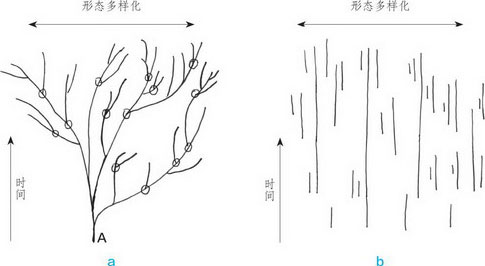
根据达尔文的演化理论,所有生物都是由同一祖先演化而来,即图中的A。经过数亿年的发展,A不断演化,发展成许多物种,隶属于不同的属、科、目、纲、门 ,甚至不同的界,最终组成一个树状的生物遗传链(图a)。但是,在这条遗传链上,每一个画圈的部分都代表一个中间类型的缺失,导致实际物种的关系是图b这种形 式,并不能组成一个生物遗传链。
直言不讳地说,如果每个地层的初期和末期之间没有这么严重的中间类型的缺失,我的学说也不会受到如此大的质疑。不过,令我没有想到的是,就算在保存得 最好的地层中,对生物变种的记录仍旧是这么稀少。