
下载掌阅APP,畅读海量书库
立即打开



 灭绝物种之间及其与现存类型之间的亲缘关系
灭绝物种之间及其与现存类型之间的亲缘关系
灭绝物种与现存物种之间的亲缘关系
灭绝物种与现存物种之间存在着怎样的亲缘关系?下面我们一起来就此进行探讨。
所有生物都根据其来源被归纳在几个大纲里,我们能清楚地解释其相互之间的关系。通常来看,
越是古老的物种与现存物种之间的亲缘关系应该越远。
但是,正如
巴克兰
 曾经说过的那样:从分类学上看,所有的灭绝物种都能和现存物种归为一个 群,或者被鉴定为与现存物种相似的类群。
曾经说过的那样:从分类学上看,所有的灭绝物种都能和现存物种归为一个 群,或者被鉴定为与现存物种相似的类群。

巴克兰
更为真实的情况是,灭绝物种常常填充了现存的属、科、目之间的缺陷类型。遗憾的是,这个观点不仅常被人们忽视,还得不到一些人的承认,于是我们有必要 探讨这个观点,并举出例子进行证明。
如果仅仅在同纲物种中探寻灭绝物种和现存物种的关系,那远不及放在整个系统中得到的结果全面。
● 欧文教授 在论文中,经常称呼灭绝动物为概括型。
● 阿加西斯 习惯在论文里称呼灭绝动物为预示型或综合型。
其实,这些词想表达的都是过渡类型或中间环节的意思。杰出的古生物学家戈德里提到,他在
阿提卡
发现的很多哺乳类的化石都处在现存属之间,极有说服力地将各个属之间的壁垒打破。
——根据居维叶的分类法,哺乳动物中
反刍类
和
厚皮类
是差异最大的两个目,但随着许多过渡型化石不断被发现,欧文只好重新整理分类系统,一部分厚皮类与反刍类一起被归到同一个亚 目下。

厚皮类
——猪和骆驼之间原本看起来差异很大,但欧文用具有中间类型意义的化石不断地将两者之间的距离拉近,使之关联。
——有蹄类,即长蹄四足兽,现在被分为
奇蹄目
和
偶蹄目
两类,它们之间是通过生长在南美洲的长颈驼联系起来的。
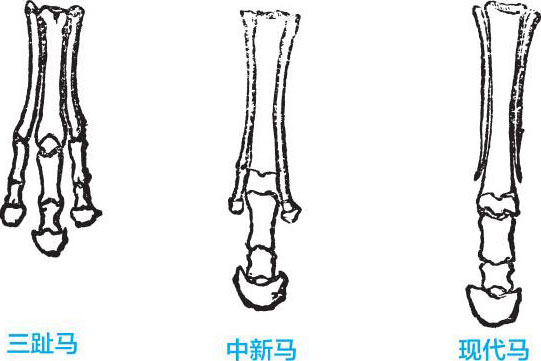
——三趾马已经被大家公认,是介于现存马类与古代有蹄类之间的过渡类型。
——在南美洲发现的,被热尔韦教授命名为印齿兽的生物非常奇特,无法被归入哺乳动物中的任何一个目。
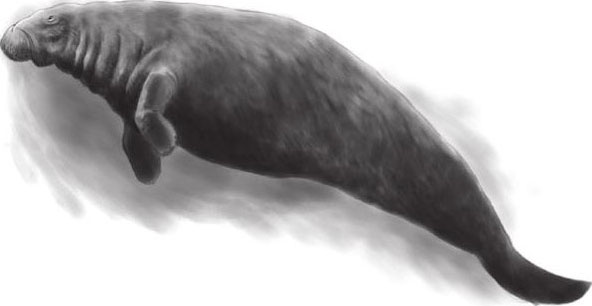
——海牛类也是哺乳动物中十分特别的一个类群,现存的儒艮和泣海牛不仅没有后肢,与后肢有关的一点儿痕迹也没留下。
三趾马 指哺乳动物纲奇蹄目马科的一类动物,体型比现代马小,前后肢均为三趾,中趾粗而着地,侧趾较小而不着地。其化石大量发现于欧洲 、亚洲、非洲和北美洲上新世地层中。我国山西、陕西、河南、新疆等地也有发现。其马趾与现代马马趾的差别,恰好印证了达尔文的进化观点。
海牛类 即海牛目(脊索动物门,哺乳纲),在海洋哺乳动物中是相当特殊的一群,因为其是由陆生转海栖的特殊种类。本目物种均为植食性, 以海草与其他水生植物为食。现存4种海牛目动物,分为两个科:海牛科及儒艮科。海牛科包含3种生存在大西洋水域的海牛;儒艮科则包含生存在太平洋及印度洋的儒 艮及已灭绝的大海牛。大海牛一直在白令海生存至18世纪才因人类猎捕而灭绝。本目已知最早的化石出现在距今约5千万年的始新世早期,渐新世与中新世是海牛目最 繁盛的时期,之后便逐渐衰退,原因可能是气候寒冷化、海洋环境变动或是古代人类猎捕造成。最早的海牛目动物大小相当于今日的猪,有四只脚,居于岸边。儒艮科 约在始新世末期出现,外形已相当接近今日的儒艮,出土化石不少;海牛科可能在渐新世初期出现,现有化石数量较少。目前对于海牛目的演化仍有许多疑问,各科间 点也尚有争议,但关于“由陆生至海栖”的演化途径由于化石证据充足已被广泛接纳。
儒艮
械齿鲸 又名龙王鲸,是已经绝种的古代海洋哺乳动物,现代鲸的近亲,生存于距今3900万~3400万年的始新世晚期,是人们已知的原始鲸类之 一。现代进化学认为,它是由陆地动物进化而来的,随后又进一步进化为两种现代鲸类。由于械齿鲸的外貌特征和巨大的海蛇相似,所以刚开始被误认巨大的海洋爬虫 类,并被命名为帝王蜥蜴。该类最早发现于美国路易斯安那州,随后古生物学家从埃及与巴基斯坦发现的化石中辨认出其他两个种。古生物学家对于它们已经退化的短 小后肢非常感兴趣,因为其可以证明现代鲸鱼原先是由陆生哺乳类动物演化而来。
——弗劳尔教授指出,灭绝的海豕拥有骨质化的大腿骨,大腿骨与骨盆内的杯状窝结合在一起形成关节,这一点看起来与有蹄类非常相似。而海牛的身体构造则 在别的地方与有蹄类相似。
——鲸鱼类和其他所有的哺乳类动物差异都非常大。发现于第三纪的械齿鲸和鲛齿鲸
被一些博物学家划分为 单独的一个目,但是赫胥黎
认为它们应该是食肉兽过渡为水栖动物的中间类型,肯定属于鲸类。

赫胥黎
鸟类与爬行类
赫胥黎曾指出,鸟类与爬行类之间虽然差异巨大,但是一些动物出人意料地将它们联系起来,这其中包括鸵鸟
和已经灭绝的始祖鸟,还包括恐龙——所有陆栖爬行动物中最大的类群——之中的细颚龙。无脊椎动物方面最具权威的学者巴兰德也认为,古生代的动物全 都能归入现存类群中。在远古时期,各个群之间的差异并没有现在这么大。
有些学者认为,并不是全部灭绝物种或群体都能作为中间类型,放到任意两个现存物种或群体之间。如果这种看法想表达的是,灭绝物种并不能作为中间类型, 具备两个现存物种之间的全部过渡性状,那么这种看法也许没有错。但是,在实际的自然分类学中,很多生物化石确实是两个物种之间的过渡类型,一些灭绝的属也将 其他现存属关联起来,甚至将一些不同的科关联起来。
细颚龙:一种体形小巧灵活的恐龙,其骨骼与始祖鸟十分接近。

鱼类与爬行类
像鱼类和爬行类这两种差异非常大的类群,假设它们之间现在有20多种差异,而在古代鱼类和爬行类之间的差异并没有20种这么多,它们之间的亲缘性比现在看 起来就近了很多。
美洲肺鱼又称星点肺鱼,隶属于脊索动物门,鱼纲,双翼肺鱼目,美洲肺鱼科。美洲肺鱼是一种古老的鱼类,现分布于南美洲静水湖泊等地,为南美洲大陆的特 产鱼类。其身体结构独特,是研究鱼类进化的重要物种。肺鱼呈鳗形,被覆埋于皮下的细小圆鳞,胸鳍、腹鳍极端退化,只留一根分节的主轴骨,呈鞭状。在所栖水域 开始干涸时,肺鱼便部分改为肺呼吸;当完全干枯时,它就钻入淤泥中,进入休眠状态,完全用肺呼吸。干旱期过后,水位恢复,肺鱼就从泥中钻出,进行繁殖。卵产 在水底挖出的穴道中,雄鱼留在其中,守候受精卵到孵化成幼鱼。在此期间,雄鱼的腹鳍肥大,生出许多具有丰富血管的丝状物,可自血液中分离氧气,有利于其幼鱼 的孵化。
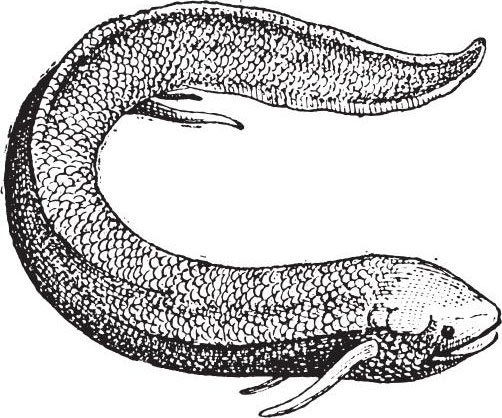
越是古老的生物类型,人们认为它身上的某些特征越能将现存生物群之间的巨大差异联系起来。 但只有变化非常大的那些类群才适用这个观点。 想要证实这个观点的正确性,也需要一些契机,因为像美洲肺鱼这样的现存物种,与几个差异较大的物种之间也有着亲缘关系。可是,当我们把古代的爬行类、两栖类 、鱼类、头足类与始新世的哺乳类做比较,或者将它们与各个类群的近代类型进行比较时,我们就会认识到这个观点的正确性了。
“物种起源”图释
关于上述事实和推论结果,我们下面具体探讨一下,它们与“物种起源”学说有多少一致的地方。
鉴于这个问题确实太过复杂,请各位看一看下面这张图:

●这里我们将标有数字的斜体字定义为属,从代表属的字母中分支出来的虚线表示该属下的不同物种。 这个图形中所画的属和种的数目之少,与 真实的自然界比起来无疑过于简化了,在此我们仅作为图例说明使用。
●图中用横线来表示连续的地层,最上面的横线用于区分现存物种和灭绝物种,横线以下的代表已经灭绝的生物类型。 由a14、q14、p143个现存 属构成一个现存的小科,b14和f14是亲缘关系很近的科和亚科,o14、i14、m14组成第三个科。这3个科与许多灭绝的属全部由祖先A繁衍而来。这些继承了祖先A身上某 些共同特征的物种,组成一个目。
第一个推导
这张图说明:
生物的性状在从亲代遗传到子代的过程中不断产生分歧,不管是什么类型的生物,相隔的代数越多,它们之间的差异就越大。所以,近代的类型与古代类 型相比较时,差异很大。
但是,遗传过程中发生的性状分歧并不是一定会出现的事,这取决于物种的后代是否能依靠这种性状分歧在自然界中占据自己的位置。所以,如同我们观察到的 一些志留纪物种的特点:某个物种在生活环境发生细微改变后,在很长一段时间内仍旧保持着与祖先相同的普遍性状。这种现象是可能存在的,图中我用F14来代表上 述这种情况。
第二个推导
与前面提到的一样,以A为祖先的各个类型,包括灭绝物种和现存物种,从属于同一个目。A目下面的物种因为遗传特性分歧不同,加上灭绝物种的影响,最终被 分为不同的科和亚科。已经灭绝的物种假设消亡于不同的年代,且还有部分物种一直生存到现在。
仔细观察这张图可以看出,倘若存在于连续地层中的多个灭绝类型,它们的祖先都是处于更深层地层中的几个点,那么它们与最上面现存科之间的差异便不会特 别大。例如,当a1、a5、a10、f8、m3、m6、m9这些灭绝的属被我们发现以后,现存的3个科之间的亲缘性便会拉近,成为一个大科。这种情况与反刍类和某些厚皮类之 间的关系极其类似。
但是,仍有一些学者认为,灭绝的属并不能填补这3个现存科之间的过渡类型,这种看法并非完全没有道理,毕竟这些过渡类型身上的特征并不能直接弥补这3个 科之间的所有差异,其中的差异可能需要好多中间型不断地进行弥补。
第三个推导
我们将某个灭绝类型所在的横线,假设成相应的地层。我们以图中的VI线为例,如果在这条横线下什么物种都没有,那么a14和b14这两个相似的科就可以合并为 一个大科,原来的3个科便重新被定义为两个科了。这两个科之间的差异也因为不断发现的化石变得更小。
第四个推导
下面,我们再来观察最上面这条线,这条线上面由a14到m148个属形成了3个现存科,假设它们之间存在6个主要特征用于相互区分,那么VI线所处的地质时期很 明显地显示出,这些物种由相同的祖先进化而来,所以它们之间的差异性会小一些,彼此之间的区别是小于6个的。我们可以看到,古代灭绝类型或多或少有一些特性 处于其变异后代之间,或者与其具有旁系亲缘关系。
总述
当然,自然环境一定比图中呈现的情况复杂多了。毕竟实际物种比图中的数量多得多,它们生存的时间也各不相同,变异的程度更不是我们可以随意想象的。
我们对地层的了解也不够全面,仅仅看到地层的最后一个部分,加之地层的记录并不是完整的,只有在极个别情况下,我们才可能将自然系统中的某个片段补充 完整,将与其相近的不同科或者目关联起来。我们只能期盼,曾经在已知地质时期出现过重大变异的类群,追溯到更老的地层时,相互之间的差异会减少一些。
所以, 同一个类群中的不同生物类型,较古老的成员之间比起现存成员之间的差异会小很多。 许多杰出的古生物学家已经发现了很多相关事实, 一致同意这种观点。
有关灭绝类型之间、灭绝类型与现存类型之间、现存类型之间的亲缘关系,借由变异的发生推导出的物种起源学说,可以对此做出圆满的解释,而其他学说都无 法对以上类型之间的关系做出合理解释。
依据物种起源学说,我们可以得知:
从一般特征上来看,历史上任何一个漫长的地质时期内,地球上的生物都是在其前后历史时期生存着的生物的中间过渡类型。
因此,在上述图中,生活在Ⅵ线(也就是第六个地质时期)的生物,是生活在Ⅴ线(第五个地质时期)生物的变异后代,同时对生活在Ⅶ线(第七个地质时期) 的生物来讲又是它们的祖先。所以,生存在第六个地质时期的生物特征,是处于第五个地质时期和第七个地质时期的生物类型之间的。
同时,我们也必须承认以下情况存在:一些古代类型已经全部灭绝了,任何区域都有可能有新物种迁入。在连续地层中漫长的间隔时期,生物体发生了许多未曾 记录下的变化。
当我们承认这些事实后,各个地层中的生物类型及其特征肯定是介于前后生物群之间的。仅举一个例子就可以说明,泥盆纪地层刚刚被发现时,古生物学家们通 过其中的化石立刻辨认出,其性状特征是介于上层石炭纪和下层志留纪之间的。但是,由于连续地层中存在不相等的时间间隔,所以每个生物类型不一定刚好看起来就 是中间类型。
整体来说,生活在各个时期的动物群,其形状特征基本上是介于前后时期的动物群之间的。 尽管有些属比较特殊,不符合这一法则,但是这种特 殊性对整个系统的影响是微乎其微的。
福克纳博士曾经按两种分类方法对柱牙象和象类动物进行排列,一种是按照亲缘关系对其进行分类,另一种是按照它们的生活年代。这样分别排列下来,得到的 结果并不相同。我们不能仅仅因为某个物种极其特殊,就认为它是最古老的物种或是新物种,具有中间类型特征的物种也不一定就存在于中间时期。
但是,因为存在很多种不同情况,例如某个物种从第一次出现到最终灭绝全部记录完整了(这只是假设,这种情况并不会出现),我们也不能就认为前后依次出 现的每个物种,其生存的时间是完全相等的。某个古老的物种有可能一直存在,比晚于它出现的物种存活的时间还长。这种情况大多出现在生存于被隔离开的海岛上的 陆生生物身上。

家鸽头部对比
我们再以家鸽为例,假如将家鸽的现存族群与灭绝族群按亲缘关系进行排列,那么排列的结果与实际物种出现的时间顺序并不相符,与实际物种灭亡的时间也是 对不上的。因为作为亲种的岩鸽一直生存到现在,而岩鸽与传书鸽之间的中间类型已经灭绝了。变异出长喙这一特征的传书鸽,比同族变异出短喙的短面翻飞鸽时间更 早一些。这个例子虽小,却将上述问题解释得很清楚。
中间类型和代表种
发现于中间地层的生物化石具有中间类型的特征,这一点在某种程度上说,与地质学有着紧密的关联性。所有古生物学者都承认,两个连续的地层之间化石的关 联性,比年代上相距较远的地层之间化石的关联性近得多。
皮克特举的这个例子,可谓众人皆知:即便不同物种之间仍旧存在差异性,但白垩纪各个地层之间的生物遗骸通常都具有相似性。就因为这个事实具有普遍意义 ,皮克特似乎不再坚信物种不变理论。但凡熟悉地球上现存物种分布的人,是不会用古代时期地球上各个区域具有相似的自然地理条件这一理由,去解释为何紧密相连 的地层中生物的差异性非常小这一问题。
至少,我们不能忽视,生活在海里的生物类型是几乎同时在地球上发生家鸽头部对比着变化的,但不同海域的气候和物理环境差异非常大。想象一下,更新世包括整个冰河时期,气候经历了那么大的变化,而当时海栖生物的变化却没有受到多少 影响。
发现于连续地层中的生物遗骸,因其各自的差异被定义为不同的物种,但是彼此间不容忽视的相似性,对于物种起源学说来讲,意义仍是十分重要的。鉴于地层 之间的沉积作用经常被间断,造成连续地层之间出现大段的时间间隔。如前面我表述过的一样,我们不能期盼在某个或者某几个地层中,找到某段历史时期中最初出现 的物种,以及全部的中间类型和最后灭绝的物种。
但是,我们在间隔期过去后能找到与之前的类型相似的物种,这些物种也被某些学者称之为代表种。正如我期盼的那样,我们不仅找到了这些相似的代表种,也 不断证实物种正以非常缓慢的速度,在人们难以察觉的情况下持续地进行着变异。