
下载掌阅APP,畅读海量书库
立即打开



在历史上首先出现农业社会的重要地区,它们的生态系统不像热带非洲那样从本质上抵制人类的改造。在温带地区,能随时利用人口增长所带来好处的潜在寄生物不仅数量少,而且也不那么可怕。但是,因为自然平衡的突破性改变都发生在5000年到1万年前,所以不可能像对非洲那样,推理出或观察到特定的农业发明和领地扩张所带来的疾病代价。
不过,我们仍旧可以推导出所有文明社会或早或迟都会遭遇的接触疾病方面的某种重要而普遍的变化,那就是,即便无须借助中间宿主,农业社会不断稠密的人口最终也会达到可以无限维持细菌和病毒感染的程度。这种情况在小型社会通常不会发生,因为与多细胞寄生物不一样,细菌和病毒的入侵会在人体内引起免疫反应,而免疫反应会要求在宿主—寄生物的关系上做出择优选择:要么受感染人迅速死亡,要么受感染人完全恢复而入侵者被驱逐出宿主的身体组织。再次感染需等到有免疫力的抗体淡出血液循环之后,那么至少需几个月或几年的时间。
不过,像生物领域里的普遍情形那样,事情绝不是一两句话所能表达清楚的。个人对感染的抵抗不只是形成抗体的问题,在某些情况下,有些病原体虽然引发抗体,但仍然可能在病人体内潜伏几年甚至一生。像著名的“伤寒玛丽”(Typhoid Mary)的携带者可以无限期地携带某种病原体,并且把可怕的甚至致命的症状传染给别人时,自身却没有明显的病症;在另外的情况下,一种传染病也可以变成隐性的病原,即潜伏于宿主身体的某些部位,并在那里长期隐藏。
最有名的一种潜伏感染模式可以让水痘病毒退到输出(efferent)神经组织中,潜伏50年之久,然后等到感染者年老时再重新发作,引发带状疱疹。这样,病毒就完美地解决了在小型社会中如何保持传染链不中断的问题:即使每一个接触到的人都感染了水痘,并且产生了免疫力,但几十年以后,当没有抗体的新一代人成长起来时,感染又会重现江湖,潜伏于该人群年长者体内的病毒就会沿着输出神经蔓延到皮肤上,产生带状疱疹;然而,一旦传染到新的宿主身上,该病毒引发的仍是习见的儿童症状,即水痘。该病对大部分人来说,症状并不严重,加上它表现出的显著的潜伏方式,都说明这是人类久已有之的病毒性传染病。在这方面水痘与现今常见的其他儿童病不同。

缺乏上述生存技巧的病菌,若又遭遇宿主体内的因抗体反应所产生的激烈取舍后果,其生存便只得依靠潜在宿主的庞大数量,也就是说,如果社会的整体规模足够大,那么总有尚未感染这种疾病而易感的人群存在。这种寄生物,若按生物进化的时间尺度来衡量,无论怎么说都只能是晚辈,尽管按人类历史的时间尺度来衡量,它已经古老得无从追溯了。只有在上千人的社会里,这种疾病才会延续,频繁的交往可以使疾病不间断地从一人传到另一人,而这类社会就是我们所谓“文明”的社会:规模巨大、组织复杂、人口密集,而且毫无例外地由城市掌管和控制。因此,直接在人类之间传播而无须通过中介宿主的细菌和病毒性疫病,首先是文明社会的疾病(即所谓“文明病”):乃城市和与城市相连的农村的特殊标识和疫病负担。它们包括麻疹、腮腺炎、百日咳、天花等,几乎是所有现代人都熟悉的常见儿童病。
儿童病花了几千年才扩散到全球,本书将用很大篇幅探讨这一扩散过程的关键性阶段。我们必须假设这些疾病(或今天已知传染病的始祖)最初的形成过程本身必定是渐进性的,包含了无数错误的开始和致命的遭遇,其中人类宿主的死亡、入侵的寄生物被消灭、传染链因此而中断,使它终未能成为文明社会的生物平衡中正常的、地方性的、或多或少带有稳定性质的因素。
在多数情况下,文明社会所特有的传染病原本都由动物传给人类。人类同驯养的动物联系紧密,因此毫不奇怪,人类常见的很多传染病与家畜(禽)的传染病存在着明显的关联。比如,麻疹可能与牛痘或犬瘟热有关;天花肯定与牛痘一类动物传染病有关;流感则是人猪共患。
根据标准手册(standard book)的记载,
今天的人类与家畜共有的疾病计有:
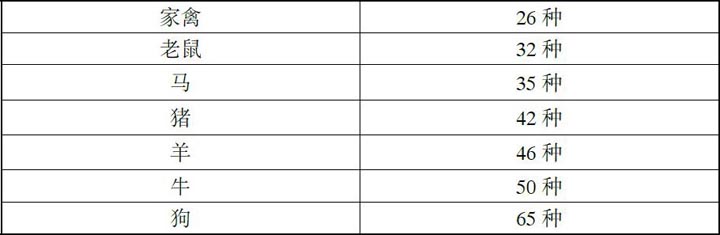
这个统计有很多重叠,某些传染病在感染人的同时还感染不止一类动物;而且,既然一些传染病很少发作,而另一些则很常见,只是统计种类就不是特别有意义。不过,大量的重叠确实表明我们同家畜(禽)的疾病联系是多么盘根错节;它还表明,随着人与动物之间亲密度的提高,共同患病的概率也在提高。
除了源自家畜或与家畜(禽)共有的疾病以外,人类也可能因卷入野生动物内部的疾病循环圈而得病。横行于穴居啮齿动物的腺鼠疫、蔓延于猴群之中的黄热病,以及蝙蝠易患的狂犬病,都属于这类较为致命的传染病。
寄生物寻找新宿主的过程还远没有结束,甚至就在近代,这一过程还造成了出人意料的严重后果。比如,牛瘟在1891年侵入非洲,杀死大量家畜以及羚羊一类的野生物种;但因它的肆虐如此严重和突然——死亡率高达90%,所以并没有在当地扎下根来,
相反,可能因为缺少易感染的有蹄类动物,它在几年后就消失了。1959年,一种叫作“奥尼欧”(Onyong nyong fever)的热病出现在乌干达——这一人类新病可能源自猴子身上的病毒。其传播速度快、范围广,但对人类的影响却比较温和,而且随着免疫力的产生恢复得很快。结果同非洲羚羊所患的牛瘟一样,奥尼欧热病也未能作为一种地方病存在下来,而是神秘地消失了,正如它神秘地出现——可能退回它原来居留的树冠区域。
10年以后,即1969年,另一种比乌干达的热病更致命的热病出现在尼日利亚,这个所谓“拉沙热”(Lassa fever)的新疫病是以医疗站里初次发现它的西医的名字而命名的。这种热病最终于1973年被证明源自啮齿动物——被认为是该病主要的宿主。于是,人们便采取了适当的预防措施来阻止该病的进一步传播。
可以想象,随着新动物的驯养、新植物的种植,以及人口的增长,这类插曲还会不时出现:传染病必定不断地从动物,尤其与人类长期紧密接触的驯养动物中传到人类身上。这种传染自然可以多向度进行,比如,有时候人的疾病也会传染给他们的家畜(禽)。同样,传染病可以在家养和野生的动物之间互换,既可以发生在同类间,也可以跨越物种界限,这是由接触机会以及潜在宿主的易感程度来决定的。
换言之,当新的生态龛由于人类活动改变了动植物的自然布局而空出后,致病寄生物在利用新的机会占领新的生态龛方面,和人类一样成功。人类的成功意味着动植物种类的减少,而每一种类数量的增多,在这一经过改进的新的饲养环境中,寄生物只要侵入单一的物种,就能够大量地滋生。几乎所有的病毒和细菌在侵入机体后,只能活跃几天或几周时间,就会被抗体阻断其在单个宿主体内的发展。
在继续探讨疾病史之前,我想还是有必要先来看看采用传染病方式的微寄生和采用军事行动的巨寄生之间的相似性。只有当文明社会的财富和技能积累到一定水平,战争和掠夺才能成为经济上可行的事业。如果武力掠夺收成导致农业劳动力很快被饿死,这还不是稳定的巨寄生模式。但这种情形经常发生,甚至可以跟1891年的非洲牛瘟感染相比,后者也是大量杀死宿主,以致未能建立起稳定持续的传染方式。
在文明历史的早期,成功的掠夺者变成了征服者,他们学会这样掠夺农民,即从后者那里抢走部分的而不是所有的收成。通过试错法可以且确实能够建立起某种平衡,生产者通过生产超过自身维生所需的谷物和其他粮食,在这样的掠夺中生存了下来。这种剩余正可以看作应付人类巨寄生的抗体,成功的政府可以使纳税人对灾难性的掠夺和外敌入侵产生免疫力,正如轻微的感染可以让它的宿主对致病的疫病产生免疫力。疾病的免疫力通过刺激抗体形成,以及将其他生理防御能力提高到更活跃的水平而发生作用;政府则通过刺激食物和原材料的更多生产以供养掌握数量庞大、武器精良的专业武士,来提升对抗外来巨寄生的“免疫力”。这两种抗体反应都会构成对宿主族群的负担,不过比起反复遭受突然而致命的灾难来说,这份负担显然要轻一些。
建立成功政府的结果,就是创建了一个与其他人类社群相比更为强大且更加可怕的社会。专业的武士几乎不费吹灰之力就可以战胜那些成年累月从事生产或寻找食物的人们。正如我们不久将看到的,一个适度感染、经地方病的病毒和细菌感染而在易感人群中形成抗体的社会,从流行病学的观点看,要比更简单、更健康的人类社群更为强大。可见,导致强大的军事和政治组织发展的巨型寄生,几乎可以与形成人体产生免疫反应的微型寄生相对应;换言之,战争和疫病的联系其实并不仅仅限于巧饰的修辞和经常伴随或尾随战争的瘟疫。
就像牛瘟和奥尼欧热病在非洲的传播那样,大部分的细菌和病毒等病原体的感染最初可能是不稳定的。我们可以想象,人类族群的人数曾多次因某些新的地方性疫病的流行而急剧削减。而人类易感宿主一而再地被消耗,又必定会不断地把入侵的病原体从早期农夫体内的“牧场”中驱逐出去。尽管如此,再次感染的基础仍然存在,因为驯化的动物极有可能已经是病毒及细菌性传染病的慢性携带者,这种传染病能够不断地骚扰人类。
从追溯牛、马、羊这类动物的野生状态中,我们也许就能看出它们被认为是这些传染病的慢性携带者的理由。在人类狩猎者多得足以影响它们的生存之前,它们是群栖的,成群游荡在欧亚大陆的草地上。作为单一物种组成的群体,它们正好提供了使细菌和病毒感染演变成地方病的条件,因为在足够大的群体里总可以找到易感染的宿主来完成传染链。畜群和寄生物之间漫长的相互进化足以形成稳定的生物平衡,一些病毒和细菌性传染病可能盛行于野生的牛、羊、马群,却不会导致严重的症状。这种传染病想必是畜群中的“儿童病”,只影响敏感的幼兽,但几乎不造成大的伤害。然而,它一旦传入人体,通常会变得很凶猛,因为初次感染的人体缺少必要的免疫力,而熟悉它们的老宿主则从一开始就至少拥有局部的防护能力。
不过,我们必须假设,在不同的地方、不同的时间,各种病毒性和细菌性寄生物最终仍成功地传给了人类,并同它们的新宿主建立了长期的关系。在很多的甚或是全部的情况下,迅速的和半灾难性的早期调适无疑是必需的,宿主和寄生物的大量死亡可能交替发生,直到新宿主发展出的免疫力和寄生物达到的适应性使传染病地方化。当今似乎已难以找到发生这一过程的例子,但澳大利亚的野兔受到恶性传染的事例表明,当病毒感染传到新群体时,它是如何生存下来并成为地方病的。
这个故事颇有戏剧性。英国殖民者在1859年把野兔引入澳大利亚,在缺少天敌的情况下,新物种迅速扩散到整个大陆,数量众多,并且对人而言已经变成了害虫:它们吃掉本应属于绵羊的草,澳大利亚的羊毛产量由此减少,无数牧场主的收益也跟着缩水。人类在澳大利亚尝试消灭野兔的努力直到1950年才出现转机,当时多发性黏液瘤(人类天花的远亲)的病毒被成功地植入澳大利亚的野兔群。最初的效力是爆炸性的:仅仅在一个季节里,相当于西欧那么大的地区就全被感染了。第一年,感染这种病毒的野兔的死亡率高达99.8%,第二年降到90%,而7年以后,死亡率仅为25%。显然,非常有力的和迅速的自然选择分别在野兔和病毒当中发生了,采自野兔身上的病毒样本,其毒性在逐年降低。尽管如此,澳大利亚野兔的数量再也没有或许永远也不可能恢复到它以前的水平——截至1965年,生活在澳大利亚的野兔只有多发性黏液瘤引进之前的五分之一左右。
1950年以前,在巴西的野兔中多发性黏液瘤是一种常见病,该病毒只在这里的野兔当中引发轻微的症状,呈现出相对稳定的地方病的发病模式。因此可能有人会认为,从巴西野兔向澳大利亚野兔的传播过程中所需要的调适,程度上应不如寄生物从不同类的宿主那儿传播到人类所需要的调适。但事实并非如此,尽管它们共有一个名字,但美洲野兔和欧洲、澳大利亚野兔并不是同种,因此,1950年在专家眼皮底下发生的宿主转移,与某疾病突破动物宿主的界限开始感染人类,从而成为人类重要疾病的可能性方式相似。
无论新疾病开始时是否像多发性黏液瘤那样致命,宿主和寄生物之间的相互调适过程本质上是一致的。一种稳定的新疾病模式,只有当双方从最初的接触当中存活下来,并且通过适当的生物的和文化的
调适达到相互容忍的关系时才算确立。在调适的全过程中,细菌和病毒拥有产生下一代所需时间比人类短得多的优势,因而有助于病原体产生在宿主间安全传播的基因突变,它要比人类遗传天赋或生理特征的相应改变快得多。的确,在后面的章节中我们将会看到,后来的历史表明,人类要将其对剧烈新疫病的反应稳定下来,大约需要120~150年的时间。
通过比较,我们看到澳大利亚的野兔数量的最低点出现在1953年,即多发性黏液瘤首次暴发的3年以后。考虑到野兔代际的短暂——澳大利亚野兔从出生到生崽只要6~10个月
,按每代人25年计算,野兔的3年相当于人类的90~150年。换言之,人类和野兔需要大致相当的代际时间来适应致命性的新疾病。
我们可以把宿主与寄生物之间相互调适的整个过程,设想成生物平衡形成之前一系列的波浪状动荡。最初的动荡可能非常剧烈,像发生于1950年的澳大利亚野兔那样,在很多情况下,由于寄生物向新宿主的转移太过剧烈而无法长久持续。然而,只要新的传染病能够无限期地生存,自然会出现动态的平衡:频繁感染期与疫病衰弱甚至几乎消失的时期交替出现。这些变动往往会形成程度不等的有规律的循环,也就是说,只要来自外部的重大入侵不改变新兴的宿主和寄生物间的平衡模式。很多的因素都会参与到这种周期性的平衡中来,比如,温度和湿度的季节变化往往使春季成为温带地区现代城市中儿童病的多发期。
人口中易感人群的数量,以及他们是群居还是散居,都是基础性的因素。例如,学校和军营一直是现代社会两个最重要的易感的年轻人的聚集场所,当代西方社会的父母都能觉察到小学在传播儿童病方面扮演的重要角色:在普及疫苗接种以前的19世纪,法国军队中的来自农村的士兵容易患病,有时非常严重,而这些传染病在城里的同龄人那里因已有过接触而具有免疫力。结果,强壮的农民子弟比那些来自城市贫民窟的营养不良的士兵死亡率更高。
感染一个新宿主所需的剂量,传染病从一个人身上传到另一个人身上所需的时间,诸如此类的传染方式,以及影响交互传染机会的习俗,都决定着多少人得病和什么时间得病。通常,只有人类宿主大量聚集在大都市,传染病才能长久地生存下去。在这里,为维持传染链不中断而与足够多的易感新宿主接触的机会,显然远多于潜在宿主稀疏分布的广大农村地区。不过,一旦乡村社会也有了足够多的易感人群,这类传染病也可能以城市为中心向外扩散,像恐怖的野火在村与村、家与家之间蔓延燃烧。然而,这类传染病的暴发,来得快,去得也快。随着当地易感宿主的难以为继,传染病也就消失了,只有它最初出现的城市中心是例外。在那里,足够多的易感人口仍然存在,使病原体不致消亡,而待到缺乏免疫力的人口再度聚集于在农村地区,又一回合的疫病暴发便再一次成为可能。
有时,所有这些复杂的因素却会沉潜下来,成为相对简单的、普遍的发病模式。对麻疹在现代城市社会中传播方式的详细统计研究表明,某种波浪式向前推进的方式在每隔不到两年的时间里达到一次高峰。而且,最近的研究发现,要维系这种模式,麻疹的持续性要求有至少7000名易感者。考虑到当今的出生率、城市的生活方式以及送孩子读书的习惯(在这里麻疹可以在初次接触该病毒的一个班级的孩子当中迅速传播),这一数字意味着,麻疹若要在现代城市中延续下去,其人口下限大约为50万。另外,通过在农村地区的散布,稍小规模的人群也可以维持麻疹的传染链不致中断。真正让该病毒难以为继的人数底线为30万~40万人。这一点,可由麻疹感染在那些人口高于或低于这个底限的海岛上的表现方式来加以证明。
但在现时流行的疾病当中,再没有别的疾病展示出如此明确的方式,可能也没有别的疾病需要如此大的人群规模来保证它的生存。对于别的常见儿童病,哪怕是相对精确的研究也尚未展开,这在很大程度上是因为人工的免疫措施在所有现代国家中已经深刻地改变了传染方式。不过,最常见的儿童病的毒性和发作频率,无论在最近还是欧洲各国政府首次开始搜集有关各类传染病发病情况的统计数字的19世纪,都出现了明显的变化。换言之,病原体和人类宿主之间的相互调适,无论过去还是现在都一直处于快速的演进之中,以应对人类生活环境的变迁。
那么,当代儿童病的“始祖”是在何时何地开始首次侵害人类的呢?为此搜寻相关的历史记载可能令人沮丧。首先,古代的医学术语很难适用于当代的疾病分类。其次,症状的变化之大,甚至已令人无法辨认。新疾病在开始的时候表现出的症状,在后来宿主逐渐产生抗体后往往就消失了。
关于以往的这类现象,有一个众所周知的例子,就是梅毒在欧洲最初暴发出来的症状。在今天,当新疾病第一次侵入到刚打破封闭的社会时,我们仍然可以观察到类似的情形。实际上,这些变化了的症状确实可以完全掩饰该疾病的性质,除非专家通过细菌学分析才能判断出来。例如,当结核病第一次进入加拿大的印第安人部落时,他们的身体器官被病菌攻击,但在白人身上却未见有病理反应。而且与那些早就接触过结核病的社群所表现的感染情形相比,不仅诸如脑膜炎之类的症状表现得更为严重,其病情的发展速度也快得多。在病症最初出现的时候,只有显微镜下的分析才能让医生确认这就是结核病。然而到第三代,随着宿主与寄生物的相互调适逐渐接近常见的城市发病方式,北美印第安人的结核病症状也倾向于集中出现在肺部了。
宿主与寄生物的调适过程是如此快速而花样繁多,以至我们必须假定现今流行的传染方式只是瘟疫当下的表现,历史已深刻地改变了它们的症状。然而,鉴于已知现代城市保持麻疹流行需要50万人,值得注意的是,最新有关古代苏美尔——这个世界上最古老的文明发源地——的总人口的估算,正好是这个数字。
似乎可以肯定,当时苏美尔诸城市之间有着足够密切的联系,足以构成一个单独的疾病库:若真的如此,接近50万的人口规模肯定足够支撑类似现代儿童病这样的传染链。在随后的几个世纪中,随着世界其他地方也开始了城市文明,连续的传染链可能也开始在别的地方出现。首先在这儿,然后在那儿,一种又一种的病原体可能就这样侵入到随处可见的人类宿主,并在由日渐增加的人口密度为它所创造出的适宜的生态龛上稳定下来。