
下载掌阅APP,畅读海量书库
立即打开




我们再对突变的其他事实和概念进行有益的探索和讨论,虽然是形式化的评论,但也不需要直接说明它们是如何从实验数据中推导出来的。
一条染色体在其自身所在区域内发生突变是可以被我们直接观察到,并得以确认的。与这条染色体的变化相比较,同源染色体的对应位置上没有发生一点变化(图8给出了示意图,×表示突变的位置),这一点是十分重要的。通常在后代中有一半的突变体性状可以直接显现出来,另一半则保持正常没有变化,这刚好证明了在突变个体与非突变个体的杂交中一条染色体受到突变的影响,而另一条染色体没有变异。这和理论的预期一样,证明了是突变体发生减数分裂时两条染色体互相分离的结果——如图9所示。这个图是一个“谱系”,一对染色体表示了三代连续的个体。如果个体的突变体的两条染色体发生突变的话,那么其子女全会得到相同的遗传性——与他们的父本、母本都不相同。
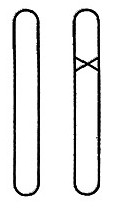
图8 杂合的突变体。

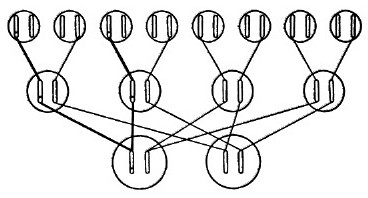
图9 突变的基因。
然而,有一个无可争议的事实:这个领域内的实验十分复杂,并不是我们前面所说的那样简单。这是由于偶然性的突变发生的时候并非是显然的,而是潜移默化的。什么意思呢?偶尔也有这样的情况出现,突变体里的两份“遗传密码本的拷贝”有些出入;至少发生突变的位置已经是两个不同的“密码”信息了。于是,有些人把原始的密码看做是“正统的”,把突变体的密码看做是“异端的”,这种认识是错误的;因为我们知道正常的突变也是从突变那里发展而来的。
在现实中,有两个版本,个体的“模式”在两者之中仿效一个。当然,这些版本可以是正常的,也可以是突变的。两者中被仿效的那个版本叫做显性,剩下的另一个版本叫做隐性。也就是说,根据模式的改变是否受到直接的突变,我们可以称之为显性突变或隐性突变。
隐性突变的概率有时甚至比显性突变要大,尽管它在突变开始发生的时候不明显,但它们的重要性是不言而喻的。只有两条染色体上都发生了隐性突变,才会影响到模式的改变(图10)。两个等同的隐性突变相互杂交或一个突变体自交时,就会产生这样的个体;在雌雄同体的植物那里这种情况是经常发生的,甚至是自发产生的。在这种情况中,可以观察到有隐性突变的模式在后代中约占四分之一。
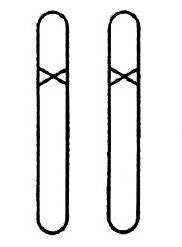
图10 纯合突变体,